 魚が陸上を歩くまで
魚が陸上を歩くまで
第2章 魚類から四肢類へ
オルドビス紀から石炭紀における魚類の系統ごとの進化の概要
以下では無顎類、板皮類、軟骨魚類、棘魚類、初期硬骨魚類、条鰭類の初期の進化を肉鰭類の進化と関連付けて概観する。
第2章-1 無顎類
無顎類の最も初期とされるコノドント網はカンブリア紀前期には出現しており、カンブリア紀中期までに、ヌタウナギ網、頭甲網、翼甲類、欠甲網などいくつかのグループを形成してオルドビス紀、シルル紀、デボン紀と繁栄した。しかし、石炭紀になると多くのグループが絶滅し、現生種としてはヌタウナギ目や頭甲網に属するヤツメウナギ目が各々数十種のこっているだけである。
最初期の無顎類は硬組織が少ないため化石として残りにくい。カンブリア紀前期、520Myaに棲息していた中国の澄江動物群のなかでハイコウイクティス:Hykouichthysは、頭甲網に属する最も初期の脊椎動物とされる(Shu et al. 1999)(Shu et al. 2003)(Degan 2003)。図1の(a)(b)に化石を、(c)に模式図を示す。体長5センチほどで、頭部には目や鼻に対応する器官や鰓を、尾部には筋節を、また尾鰭のほかに背鰭を持っており、すでに魚類の原型が見られる。

a. 化石全体図;左側が頭部、尾部先端部は恐らく腐敗により消失。スケール線は5ミリ。
b. 頭部拡大図;鰓や恐らく頭蓋内骨格、心臓部が見て取れる。スケール線は5ミリ。
c. 全体模式図;数字1~6:鰓。?A~?C:確かではないが、内2箇所は心臓部に対応している。 From Zhu 1999
頭甲網では体の詳細が判明できる保存状態の好い化石の多くは頭部や胸部が皮骨質の装甲で覆われている。ハイコウイクティスより後のオルドビス紀中期に棲息していたアランダスピス科の1種サカバンバスピス:Sacabambaspisは尾鰭を持つが背鰭は無く、現生のおたまじゃくしに似た遊泳をしていたとされる。おたまじゃくしよりも体部に比して尾鰭が小さいことから、遊泳能力はかなり劣っており、海底付近をゆっくり移動していたと思われる。口は腹側に付いており、その位置と形状から海底の有機物を含む泥を吸引し濾し取る摂食方式をとっていたとされる。また、同じくデボン紀に棲息していた骨甲目のセファラスピス:Cephalaspis も海底域を生活圏としていたとされるが、背鰭を持ち、対になった胸鰭もあり、アランダスピスよりは移動能力が高かったことを予想させる。
この頃の無顎類には胸鰭を持つものと持たないものが混在しており、胸鰭の有無が必ずしも進化の過程を示しているわけではない。四肢類の四肢の原型は対鰭である胸鰭と腹鰭であるが、対鰭である胸鰭を持った最初の無顎類はオルドビス紀末に現れた体長15から20センチほどの欠甲目のアナスピダ:Anaspida(Coates 2003)とされる。アナスピダは第1章の図4の3番に見るように背鰭はあまり発達していないが、逆歪尾(ぎゃくわいび)の発達した尾鰭を持っている。逆歪尾は前上方への推進力を得るのに都合の好い形状とされる。アランダスピスやセファラスピスに比べると格段に遊泳力が向上していたと考えられ、移動しながらプランクトン類を濾し分ける遊泳性の生活をしていたと考えられる。しかし、この対鰭はほとんど動かせなかったようで、遊泳時における安定化の機能を持っていたであろうが、それ以上の機能はまだ無く、せいぜい若干の昇降舵的機能を持っていた位であろう。
なお、逆歪尾は前上方への、歪尾は前下方への推進力を得るのに適した形状である。現生条鰭類の多くは前方への推進力を得るのに適した尾鰭の上下がほぼ対称な正尾を持つ。
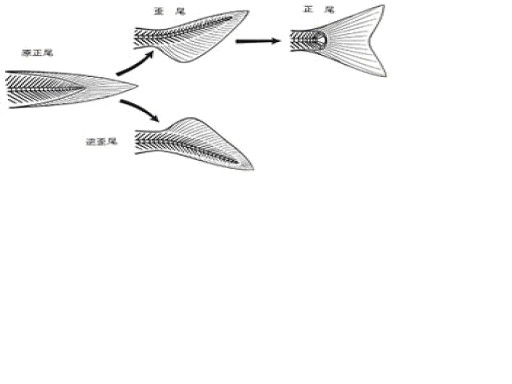
無顎類はほとんどが吸引式の摂食を行っていたが、その口の位置や尾鰭、対鰭、体型などから、頭甲類は海底泥中における底生型の、翼甲類や骨甲類は海底近くを移動する半遊泳型の底生、そして欠甲類は遊泳型の生活方式を主に採用し、互いに棲み分けていたと思われる。
無顎類が出現した当時海サソリなどごく一部を除くと強力な捕食者はほとんど存在していなかったと考えられる。しかし、無顎類の多くは甲皮魚類、あるいは甲冑魚と呼ばれるように頑丈な外皮で頭部や胸部が覆われており、外皮の機能に関する議論が為されている。ローマーは海サソリからの防御のためと考えた。しかし、アランダスピス:Arandaspisやセファラスピス:Cephalaspis に見られるように体の前方は硬い外皮で保護されているが、体の後方は甲皮が無いものが多い。しかも巨大な海サソリは肉食性であったが敏捷性に欠けており、少なくとも翼甲類や欠甲類のような遊泳力のある魚類の脅威にはならなかったとも考えられる。そのためベリル(「脊椎動物の起源」)は、新たに「過剰になった血液中のリン酸カルシュウムを処理するために体表に沈着させた」という説を提案した。即ち、甲皮魚類が棲息していた河口近くはリン酸塩の濃度が海よりも相対的に高く、そのため血液中のリン酸塩の濃度が高まり、一方腎臓の機能がまだ不十分だったので血液から直接リン酸塩を排出せざるを得なかったであろう。沈着は血液の流れが小さい部位ほど顕著になると考えられ、移動に支障を来たさない頭胸部がその部位になった、と考えた。
しかし、ほぼ同時期、同場所に生息していた遊泳性の欠甲類は頭胸部に装甲を発達させていず、装甲が底生の無顎類に顕著なことから、このような甲皮は底生生活に必要な機能を持っていたと考えられる。即ち、泥中から有機物に富む餌を探り当てたり、積雪をかき分けて進むラッセル車のように泥中を移動するときの除泥機能を持っていた可能性がある。また、尾鰭部の推進機能保持のために装甲が体部後方に形成されなかったと考えられる。
第2章-2 板皮類
最初に出現した顎口類、あるいは有顎類は板皮類であり、無顎類のThelodonti類から板皮類へと進化するときに顎を獲得したと考えられている(Fraser et al. 2009)。無顎類出現後30Myほど経たオルドビス紀後期、遅くとも444Myaまでには出現していたとされる(Xiaobo et al 2010)。顎を持っていたと見られるSkiichthysはその頃の魚類であり、板皮類とされるが確定的ではない。板皮類の明確な化石記録にはシルル紀初期439MyaのAntiarchiやPetalichthyida (Long 2011)、さらにより新しいシルル紀後期420MyaのSilurolepis (Zhang et al. 2010)がある。また、顎を持ったにもかかわらず吸引式の摂食を行う板皮類も棲息していた(Carr et al. 2009)。板皮類は淡水性もいたが、主に海礁を生息場所とした底性の魚類で、デボン紀初期になって急激に放散し繁栄したが、デボン紀末にはほとんど滅亡した。遊泳力がそれほど無かったため遠方との交雑が少なく地域ごとに特徴のある多様な形態を示し、7目240種ほどが報告されている。但し、ゴンドワナ大陸とローレンシア大陸が近づいたデボン紀後期には両大陸間でも交雑があったとされる(Young 2010)。
頭胸部は頑強な板状の外骨格で覆われており、これら板状外皮の間は関節されており、ある程度の可動性があった。特に頭甲と胸甲の間の可動性は大きく、上下の顎の関節にもなっていて、口は大きな開口度を持つことができるようになっている。底生で待伏せ方の捕食者であり、餌が近づくと周囲の水ごと吸い込む方式を採用していたと考えられる。消化管部に多数の小石が散在する板皮類化石が見つかっており、摂食の際に一緒に飲み込まれたと考えられる(Long 2011)。頭胸部に頑強な外骨格を持つが体の後方に装甲を持っていないのは、岩礁性乃至は砂地の環境で待伏せ型の摂食を行うのに適応したためと考えられる。即ち、頭胸部以外は岩礁の隙間や砂地で保護し、上位捕食者である海サソリ等から身を守りながら、無顎類や他の板皮類、棘魚類を捕食していたことを予想させる。
板皮類は肉食動物として位置づけられるが、顎にはデボン紀後期に出現した一部の板皮類を除き歯は無いとする説が一般的である(Smith & Johason. 2003)(Borrow. 2003)。最近シンクロトロンx線トポグラフによる詳細観察に基づき板皮類も歯を持っていたとする説が出された(Ruklin et al. 2012)。そして顎口類出現直後に顎に引き続いた歯が形成されたと結論している。しかし、調査対象の板皮類はデボン紀のものであり、440Myaには板皮類が出現していたと考えられ、初期の板皮類が歯を持っていたとは言い切れない。
腹鰭を持たない種類もかなりあるが、多くは胸鰭を持ち、肩帯の骨格と筋肉は硬骨魚類並みに発達している。胸鰭を使った方向や位置の制御は底性活動に必要な機能であった。しかし、胸鰭は外骨格で覆われており、可動性はあまりなかったであろう。板皮類はいずれも下方への推進力を生み出す歪尾を持っており、底性生活に適応していたことをうかがわせる。肺乃至は鰾は持っていないとされるが、デボン紀の後半には肺様組織を持っていた種類も出現した可能性がある(Perry et al. 2001)。体長は1メートル以下のものが多いが、デボン紀後期にはディニクティスのように10メートルにも達する巨大魚も出現している。
板皮類はデボン紀末に絶滅しているので、デボン紀中期には後述する底生へと進出した肉鰭類と生態系で競合した可能性もある。しかし、体形の大きな違いから、板皮類は岩礁や砂地に、肉鰭類は半遊泳型の底生、さらには三角州や潟といった浅い水域に棲み分けていたと考えられる。
板皮類はデボン紀末に絶滅していることもあり、板皮類と軟骨魚類の分岐年代はもとより分類上の相関も必ずしも明らかではなく、板皮類の系統樹上の位置づけに関しては長いこと議論されてきている(Trinajstic et al. 2007)。板皮類と軟骨魚類の類縁性が主張されたり、硬骨魚類と類縁とする説、総ての顎口類の祖先とする説、さらには板皮類は顎口類の祖先系と単系統系の2系統に分離できるとする説も提案されている。単系統であり他の顎口類と姉妹群の関係にあるとする説が有力視されてきたが(Carr et al. 2009)、最近の報告では軟骨魚類との類縁性が強調されているように見える。現生サメ類によく見られる胎生の板皮類の存在が確認されたり(Long et al. 2008)(Long et al. 2009) (Ahlberg 2009).(Ahlberg et al. 2009)、eyestalkの構造から板皮類と軟骨魚類との類縁性が示されている(Young 2008)。
一方、眼球を動かす筋肉系は板皮類ではヤツメナギに似ていることが明らかにされた(Young 2008)。しかし、頭蓋内痕跡化石により、板皮類の眼球運動神経は無顎類に比べると格段に長くなっており、神経内の伝達速度が速くなっていること、即ち板皮類において神経にミエリン:myelin鞘が形成されたことを示唆している(Zalc et al. 2008)。このことから、捕食のための急激な顎の開閉などの運動機能は無顎類に比べて向上していたことが予想される。
さらに最近では軟組織が保存された板皮類の化石から、板皮類の体幹には有顎類、特に魚類に特徴的な水平中隔が無いことが明らかになった(Trinajstic et al. 2007)。水平中隔は軟骨魚類を含めた魚類一般に見られる組織であり、筋肉系、脊椎系、皮膚などと機能的に結合していて、遊泳移動に関係した組織である(Westneat et al. 1993)。しかし、ヤツメウナギなどの現生円口類は水平中隔を持っていない。即ち、板皮類は水平中隔膜が無いことから、移動に関係する器官は軟骨魚類よりも無顎類に近いと考えられる。
これらの結果から、板皮類が先に無顎類から分岐し、その後に軟骨魚類が分岐したとする説は妥当性があると思われる。しかし、両者がほぼ同時期に分岐し、各々異なるニッチに進出・適応した可能性もある。底生に向かったのが板皮類へと、遊泳性に向かったのが軟骨魚類へと進化したことになる。板皮類につながる現生種が生存していないため遺伝子解析は出来ず、現時点では分岐の経緯に決着をつけるのは困難であり、さらなる化石の蓄積と詳細な解析が必要である。
なお、デボン紀後期392Myaに出現したEastmanosteusは歯を獲得したが、これは他の顎口類の歯とは独立に進化したものとされる(Smith & Johanson 2003)。また、同じくデボン紀後期387Myaに棲息していたBothriolepisは肺に似た器官を持っていたとされ(Perry et al. 2001)、もし肺ないしはそれに似た器官であれば、硬骨魚類の肺や鰾とは独立に平行進化したものと考えられる。
第2章-3 軟骨魚類
もうひとつの初期顎口類とされる軟骨魚類は化石として残りにくいが、鱗の化石は既にオルドビス紀後期の455Myaの地層から出土している(Cole & Currie 2007)(Janvier 1996)(Venkatesh et al. 2007)。しかし、歯の化石はシルル紀末期の418Mya(Miller et al. 2003)までは見つかっていず、初期の軟骨魚類は板皮類と同じように歯を保持していなかった可能性がある。体化石として現在最も古いのは409MyaのDoliodusである(Miller et al. 2003)。
分子時計に基づいた解析では軟骨魚類の分岐年代として528Myaや500Myaといった値が得られており、カンブリア紀に既に分岐したともいわれている(Hedges & Kumar 2003)(Kumer & Hedges 1998)(Inoue et al. 2003)。こういった計算では概して化石記録よりも古い分岐年代が算定される傾向があるが、鱗や歯の化石の存在もあり、遅くともオルドビス紀後期の455Myaには既に軟骨魚類が棲息していたとすることは妥当性があると思われる(Smith et al. 2002)。しかし、放散を開始したのはデボン紀中期からであり、デボン紀後期までには80種以上になった。軟骨魚類の多くは海生であったが、河口から淡水域に棲息する軟骨魚類、Doliodusも報告されている。
軟骨魚の体の形態が判別できる最も古い化石はデボン紀初期394MyaのSeretolepisで、鱗や棘を持った鰭と対鰭などが見られる(Maisey & Anderson 2001)。初期軟骨魚類がfin-spins や歯、鱗といった骨質器官を形成する能力を持っていたにもかかわらず硬骨魚類への道はたどらず、その後現在に到るまで3億年以上も軟骨魚として留まった。
特に繁栄したのが体長2メートル程度のクラドセラケ:Cladoselacheである。体型は現生サメ類とあまり変わらず、当時既に完成された遊泳性能を持つ優れた捕食者としての地位を占めていた。しかし、現生種と異なり、胸鰭や腹鰭の根元が広くなって体についているので対鰭の可動性は低かったようであるが、安定板以外にも昇降舵やブレーキの機能を持っていたと考えられる。軟骨魚として留まった理由のひとつは体形デザインの先行性にあると思われるが、3億年以上に渡り海の食物連鎖で上位位置を保持してきたことは進化の観点で興味が持たれる。
南アフリカから出土した初期軟骨魚の脳函は基硬骨魚類:stem ostheichthyanの特徴とされる耳孔や背側頭部泉門:posterior dorsal foutanelle、口蓋基盤顎関節:palatobasal jaw articulationを持っているとの報告もある(Maisey & Anderson 2001)。軟骨魚類においても出現初期にはいくつかの系統の特徴がモザイク状になっていたと思われる。
こういった混在した特徴に加えて中間化石がみつかっていないこともあり、軟骨魚類の系統樹における位置づけはまだ確実ではない。板皮類のような装甲を発達させた魚類から進化したとする説、無顎類のThelodontsと似た尾の形や鱗を持っていることから板皮類と同じ頃に無顎類から分岐したという説、総ての顎口類の祖先系であるとする説などがある。最近でも議論は続いており(Dosay-Akbulut 2006)、従来棘魚類に分類されていたPtomacanthusの脳函の詳細から、これがサメにより近く軟骨魚類乃至は基顎口類であり、サメは初期棘魚類の中から分岐したものであり、棘魚類は板皮類様の祖先から出たとしている(Brazeau 2009)。
系統樹上での位置づけがはっきりしないにせよ、板皮類と軟骨魚類は、両者の体型の違いから棲み分けていたことは確かであると思われる。即ち、前者はbottom-feeder として、後者はopen-water swimmer として生態系の位置を占めていたと考えられる。板皮類は底生型のため地域に特有の種が多いが、軟骨魚類は遊泳型なのでシルル紀前期の段階で既に世界中に棲息範囲を広げていた(Smith et al. 2002)。
第2章-4 棘魚類
初期顎口類としてさらに棘魚類:Acanthodiansがオルドビス紀後期には出現している(Long 2011)。シルル紀前期には既に世界各地に棲息しており、遊泳性に優れていたと考えられる(Smith et al. 2002)。棘魚類は同じ遊泳性でも軟骨魚類に比べると現生の遊泳性硬骨魚類に近い体型を持っている。初期には海水域に棲息していたが、条鰭類が放散し始めた390Myaころからデボン紀後期には汽水域や淡水域に棲み分け、主に淡水域で繁栄した。石炭紀後期には条鰭類も淡水域に進出したが、同じ淡水の環境で棘魚類は50Myというかなり長期にわたり共存し、ペルム紀末頃に絶滅している。何ゆえ条鰭類と長期にわたり淡水域で共存できたのかは、進化の観点から興味が持てる。遊泳力の優位性といった条鰭類の特性は、広い海域では有効であったが、浅くて狭い河川や湖といった水域ではあまり有利性が無かったのかもしれない。
棘魚類は出現後主な棲息域を海から淡水に変えてペルム紀末までおよそ170Myもの長期にわたり生存していた。その間デボン紀初期に体長2メートルのXylacanthusやデボン紀中期に体長1.5メートルのGyracanthidsなども現れたが、多くは小型のままに止まり、3目程度の主要なグループを生み出したに留まっている。繁栄したとされる淡水域の生態系でも高位の位置にはなれなかったようである。
棘魚類の典型例Acanthodiansを第1章の図4の10番に示す。対鰭の胸鰭と腹鰭、前後二つの背鰭と尻鰭を持ち、これら鰭は基本的には棘を持っている。腹鰭を1対ばかりでなく複数対を持つ種類もいた。これらの鰭は棘があるため、条鰭類の鰭より可動性が無く微妙な動きも出来なかったと考えられる。頭部は短く、先端部に大きな目を持っている。棘魚類のなかでも最も繁栄し、ペルム紀まで生存したAcanthodiformesの多くは歯を持たず、よく発達した鰓耙(gill raker:さいは:鰓の内縁の突起)を持っており、filter-feeding 方式で摂食していた。また、呼吸のための初歩的な肺を持っていたとされる。Acanthodiformesに属するHowittacanthusは現生硬骨魚類や四肢類と同様3種の耳石を持っており(Janvier 2011)、機敏な運動能力を持っていたことをうかがわせる。
棘魚類の鱗の成長に関して二つの異なる報告がある。一つは中国のシルル紀地層から出土した最初期の棘魚とされるShinacanthusであり、その鱗は生涯にわたり成長していたことを示しており、この特徴は板皮類や初期硬骨魚類にも見られる。なお、現生の硬骨魚類の鱗は、数が決まっていて成長に伴い大きくなる。一方、初期の棘魚類とされるクリマチウスは細かい鱗を持っており、軟骨魚類のように成長とともにその数を増していったとされる。
棘魚類の棘は捕食者に対する警告のためとされているが、中国のシルル紀後期の地層から出土したYealepisやカナダのデボン紀初期の地層から出土したPaucicanthusの棘は顕著ではない。特に当時のサメに見られるように根元が広がった胸鰭を持つ棘魚でこの傾向が強い(Long 2011)。初期の棘魚の一部に見られる棘の消失・減少と鰭形状の変化は、棘が運動機能にもかかわっている可能性を示唆しているように思われる。
棘魚類も系統樹上の位置づけははっきりしていない。かっては軟骨魚類と共通する多くの特徴を持っており、サメと近縁とする説が強かったが、ローマーは硬骨魚類に近縁とし、コルバートは単系であるとしていた。また、棘魚類は板皮類に近縁とする説や、軟骨魚が起源であり、硬骨魚類と姉妹群であるか、または硬骨魚類に近縁の群から分岐したという説も出されている(Brazeau 2009)。
しかし、最近になって、Acanthodiansのサメに似た特徴は、初期顎口類の多くに見られる一般的な特徴であり、特定の顎口類と近縁であることを示しているとは限らないとする解釈が出てきている。最近では保存状態のよい棘魚類の頭部骨格化石から基硬骨魚類:stem ostheichthyanの可能性が強まっている(Brazeau 2009)。サメとの類似性を除くと、もっとも繁栄したAcanthodiansは硬骨魚と共通するいくつかの特徴を持っている。即ち、棘魚類の脳函は硬骨魚類のそれと似た形状をしており、鰓皮輻(さいひふく:硬骨魚の鰓蓋に続く膜に並ぶ放射状の骨):branchiostegal raysを持ち、脳函内に軟骨質性硬骨の存在(多分)が見られ、歯が顎に固定されている。さらに3個の耳骨を持ち、神経頭蓋の耳殻部と後頭部を分離する耳―後頭裂溝(otico-occipital fissure)があり、神経頭蓋腹面に通気溝:spiracular grooveがある。こういった化石の詳細調査から、棘魚類と硬骨魚類をまとめて真口類:Teleostomiに分類することもある。しかし、棘魚類は単系で子孫を残さず中生代に滅亡したとする説もあり、現生魚類との関係においても依然として議論が継続中である。
板皮類や軟骨魚類、棘魚類はオルドビス紀後期には出現していたが、顎を獲得したにもかかわらず優位な捕食者として一気に放散することは無く、オルドビス紀、シルル紀を通じて無顎類が脊椎動物の主要な地位を占めていた。次に述べる硬骨魚類を含め、これら顎口類が放散を始めるのはシルル紀後期になってからであり、生態系で主要な位置を占めるようになるのはデボン紀に入ってからである。
第2章-5 硬骨魚類
顎口類の主役とも言うべき硬骨魚類は、鰭内に骨と筋肉を持つ肉鰭類と現生の普通の魚に見られるように鰭条のある鰭を持つ条鰭類に分けられる。両者は420Mya以前に既に分岐していたことが、化石記録と分子時計に基づく計算とから明らかにされている (Zhu et al. 2009) (Thatcher et al. 2008)。なお分子時計に基づいて計算では、肉鰭類と条鰭類の分岐時期は450Myaと見積もられている(Hedges & Kumar 2003)。ここでは肉鰭類と条鰭類に分岐する前の硬骨魚類を初期硬骨魚類と呼ぶことにする。即ち、初期硬骨魚類がシルル紀後半に肉鰭類と条鰭類に分岐し、前者の一部から後に陸上生活に適応した四肢類が出現し、後者から現生魚類のほとんどが出現したことになる。
初期硬骨魚類とされる化石は少ないが、Lophosteus , Andreolepis, Nacilepis, Orvikuina, Terenolepis, Dialipina, Ligulapepis等の報告がある(Friedman & Braseau 2010)。これらの中で比較的詳細に調査されているのはシルル紀後期の423~421Myaに棲息していたLophosteusとAndreolepisである。これ以外の初期硬骨魚類の化石記録は断片的なものしか報告されていない(Zhu et al. 2009)。
LophosteusとAndreolepisは、主に北欧から出土しており、海乃至は海岸に棲息していた(Marss 2001)。Andreolepisの復元図を第1章の図4の11番に示す。これら初期硬骨魚類が硬骨魚類の系統樹の根元近くにあるのは確かであるが、肉鰭類に近いのか、条鰭類に近いのかで議論されてきた(Lee & Kimelman 2002)(Marss 2001)。
当初LophosteusとAndreolepisは近縁であり、歯の形状や肩帯、鱗の形などからともに条鰭類の系統と考えられた。しかし、その後の調査により、条鰭類に特有の鱗のガノイン構造が前者では無く、側線の構造や鱗の分布や詳細形状にも違いが見られ、それほど近縁ではないとされる(Lee & Kimelman 2002)。Andreolepisの方がやや条鰭類に近い(Marss 2001)、乃至は棘魚類に近いとされる(Friedman & Braseau 2010)。また、Lophosteusは肉鰭類に近いが分類不明とされる(Friedman & Braseau 2010)。いずれにせよLophosteus とAndreolepisは肉鰭類と条鰭類が分岐する前の初期硬骨魚類と考えられている(Lee & Kimelman 2002 )(Esbaugh & Tufts 2006)(Botella et al. 2007)(Zhu et al. 2009)(Friedman & Brazeau 2010)。
最近の初期硬骨魚類に関する詳細報告によると、Andreolepisが最も初期の硬骨魚であり、LigulapeoisやDialipinaが、肉鰭類と条鰭類が分岐する直前の初期硬骨魚類とされる(Friedman & Brazeau 2010)。但し、LigulapeoisやDialipinaは初期の条鰭類とする見解もある(Zhu et al. 2006)(Botella et al. 2007)(Esbaugh & Tufts 2006)。
これら最も初期とされるAndreolepisをはじめ多くの初期硬骨魚類の体表は小さな鱗で覆われており、板皮類のような装甲を持たず、体型は流線型である。口吻は前方を向いており、歯は硬骨魚類や四肢類の歯に近いサイズを持っている。従って、板皮類のように待伏せ型で周囲の水ごと餌を飲み込む吸い込み式摂食とは異なり、噛み付き式摂食を行っていた捕食者であったと思われる(Bottela et al. 2007)。これらのことから初期硬骨魚類の多くは、底生型ではなく遊泳型の魚類であったとされる(Smith et al. 2002)(Cloutier & Arratia 2004)。しかも最も初期の硬骨魚類とされるAndreolepisの段階でも遊泳性魚類としては洗練された流線型の形態を示している。
遊泳性であった初期硬骨魚類は、同じ頃既に棲息していた遊泳性の軟骨魚類や棘魚類と競合関係にあったと考えられる。一方、現生肉鰭類や四肢類が肺を持ち、現生条鰭類が肺から進化した鰾を持つことから、肉鰭類と条鰭類の共通祖先と位置づけられる初期硬骨魚類は肺を持っていたことになる。
さらに最近の棘魚類化石の詳細調査から、棘魚類も初期的ではあるが肺を持っていたことが明らかにされた(松本 2006)。そのため棘魚類と硬骨魚類の共通祖先として真口類:Perfect mouth:Teleostomiの存在が考えられ、真口類の時期に肺を獲得したことになる。
分岐後シルル紀末の420Myaからデボン紀初期の410Myaにかけて種々の硬骨魚類が急激に現れ始める。シルル紀後期の419MyaにはGuiyuが、417MyaにはPsarolepisが(Sonkoly et al. 2007)、414MyaにはAchoaniaが(Thatcher et al. 2008)(Zhu & Yu 2009)、413MyaにはOnichodont(Johanson et al. 2006)が出現している。これらはいずれも肉鰭類に分類されているが、軟骨魚類の特徴も持つ。また、ほぼ同じ時期の417Myaにはハイギョと四肢類の共通祖先と考えられる肉鰭類Styloichthys(Miller et al. 2003)(Zu & Yu 2002)や411Myaにはハイギョ類に分類されるPorolepis(Cloutier & Arratia 2004)に引き続いて410MyaにはYongolepis(Friedman & Blom 2006)やPowichtys(Zhu & Schultze 1997)が出現している。また、最初期のシーラカンスとされるCrosspterygiiが418Myaに(Brazeau 2009)(Young 2010)、Onychodont が413Myaに、Eoactinistiaが408Myaに現れている(Johanson et al. 2006)。
初期の肉鰭類とされるGuiyuは体長28センチ程度で、胸鰭と腹鰭の対鰭と背鰭は棘を持ち、背鰭のほかに中間背板:median dorsal plateを持っており、板皮類に似ている。脳函が関節していることは肉鰭類の特徴である。PsarolepisやAchoaniaもGuiyuに似ているが、外皮や歯、顎の形状等に違いがある。しかし、脳函が関節を持つという肉鰭類の特徴を備えている。Styloichthysに関しては、最近の歯などを含めた詳細報告ではシーラカンスとの近縁性が見られ、シーラカンスと姉妹群とされているようである(Friedman 2007)。ハイギョの系統とされるYongolepisやPowichtysの脳函は、他の肉鰭類のように完全に分離して関節しているのではなく、部分的につながっており、より古い形を残していると考えられる。
肉鰭類は420Myaから410Myaにかけて急激な多様化を起こしており、まさにこの時期が肉鰭類の適応・放散の時代でもあった。一方、条鰭類の放散は肉鰭類よりも20My以上遅れ400Mya頃から、さらに本格的な放散はデボン紀中期390Mya頃になってからである(Long 2011)(Yu et al. 2010)。
これらの硬骨魚類に共通する特徴は、硬骨化した内骨格と顎、および歯を持っていることである。しかし、初期硬骨魚類を含めてこの頃の硬骨魚類は骨化がまだ充分ではなく、一部の内骨格が骨化している段階である。また、肩帯も胸鰭の筋肉を支持するために強化され、さらに鱗同士の重複度も大きく、これにより体幹の剛性が増し、遊泳力が向上したと考えられる。また、鰓弓にも他の顎口類に見られないいくつかの特徴があるほか、鰓や消化器の機能に関連する各種筋肉も発達している。しかし、420~400Myaにかけて出現した硬骨魚類の多くは軟骨魚類や板皮類の特徴も兼ね備えており、また、肉鰭類、条鰭類の区別も明確でないものも多い。
第2章-6 肉鰭類・四肢類
420Mya頃に急激に多様化した初期の肉鰭類は洗練された流線型の体型を示している。また、この時代の肉鰭類の口は前向きである。最も初期の肉鰭類とされるGuiyuの模式図と詳細化石図を図3に示す。非常に洗練された魚体形を持ち、口吻、鰭、鱗、体形などから遊泳性であったことがうかがえる(Zhu et al. 2009)(Coates 2009)(Qiao & Zhu 2010)。
さらに現生シーラカンスの観察から明らかになったように、シーラカンスは肉鰭を水底を這うのに使うのではなく泳ぐのに使用している。シーラカンスは肉鰭類が分岐した直後に出現しており、従って初期の肉鰭類の多くは底生の環境に適応していたbottom dwellerではなく、対鰭を泳ぐのに使用する初期硬骨魚類の特徴を引き継いで、遊泳性の生態系に適応していたことを支持すると考えられる。

a. 復元図; 灰色部は不明で、他の初期硬骨魚類から類推 b. 化石対応図;ano:前鼻孔
br:鰓条骨 cla:鎖骨 cle:cleithrum drs:背鰭鱗 dsp:背鰭棘 et:外側頭骨(extratemporal) eta:付属外側頭骨(accessory extratemporal) f.add:内転筋窩 f.gl:関節窩 gu:咽頭 ju:頬骨 l.ext:側外肩甲骨(lateral extrascapular) lj:下顎 m.ext:中央外肩甲骨(midean extrascapular) mx:上顎骨 n.sp:通気孔切り欠 op:鰓蓋 pa:頭頂骨盾 pcl:後擬鎖骨(postcleithrum) pop:前鰓蓋 ppa:後頭頂骨盾 psc:前上擬鎖骨(presupracleithrum) pt:後側頭骨 scl:上擬鎖骨(supracleithrum) sop:補助鰓蓋 sp:胸部脊椎 tr:鱗状鰭条(lepidotrichia) vrs:腹部鱗 From Zhu et al. 2009
初期の肉鰭類に分類されるGuiyu, Psarolepis, Achoaniaが419~414Myaに出現したのに引き続いて、ハイギョの系統とされるStyloichthys, Onycodont, Porolepis, Yungolepis, Powichthys が416~410Myaにかけて、さらにシーラカンス系とされるEoactinistiaが408Myaに相次いで出現している。即ち、420Myaから400Myaにかけては肉鰭類の放散の時期であったが、これらハイギョの系統とシーラカンスの系統を除くと、後の脊椎動物の上陸につながる肉鰭類は414MyaのAchoania以降395Myaに出現したKenichthys(Zhu & Ahlberg 2004)(Janvier 2004)までの19Myの間あまり化石記録がないようである。
Kenichtysは、後の陸上での呼吸と関係する内鼻孔を持っていることが確認された最初の肉鰭類である(Janvier 2004)。この内鼻孔は歯列を横断する位置にあり、元来は前外鼻孔と後外鼻孔の一対であった鼻孔のうち、後外鼻孔が、元の位置から後方に移動して前上顎骨と上顎骨を隔てる歯列を横断する位置に移動したことを示している。これにより長い間議論されてきた内鼻孔の起源と形成過程が明らかになった。即ち、現生陸棲脊椎動物の内鼻孔は後外鼻孔が口蓋へ移動し、口の中を徐々に喉側に移動したものである(Zhu & Ahlberg 2004)。
Kenichtysは395Ma頃に棲息していたとされるが、内鼻孔を持っていたことは底生であったことをうかがわせる。即ち、水棲の嗅覚器官としての鼻孔は、前外鼻孔から後外鼻孔へと水が通り抜けるときに機能する。遊泳型であれば移動に伴い鼻孔内で自動的に水流が生じるが、底生になると鼻孔内水流を自発的に引き起こす必要がある。そのために後外鼻孔を口蓋に移し、口や鰓を動かすことで鼻孔内に水流が生じるようにしたと考えられる。即ち、395Mya頃に一部の肉鰭類は遊泳型から底生型へと棲息域を変えたと思われる。脳函は肉鰭類の特徴とされる間を蝶番で結合された2分割になっており、急激に口を大きく開くことが出来るようになっており、周囲の水ごと餌を口中に取り込む丸呑み方式に適した形態であったと考えられる。
現生シーラカンスとハイギョは内鼻孔を持たないが、Kenichtysが出現するより前の遊泳型肉鰭類の系統から分岐したためと考えられ、上記仮説、内鼻孔を持つKenichtysが底生であったことを支持すると思われる。
デボン紀後期に起こった最初の大絶滅事変、Kellwasser eventをはさんで385Myaから365Myaにかけて底生型肉鰭類の一部はさらに潟、三角州といった潮汐域に進出した。385Mya頃棲息していたEustenoptelonの対鰭骨格や対鰭帯には四肢の骨格の原型が見られるので、この頃に潟、三角州といった潮汐域に進出したと思われる。肉鰭類に特有のコズミン鱗は380Myaころ棲息していたEustenoptelonあたりまでは保持されているが、体表の皮骨はエナメル、あるいはエナメロイドが消失しており、軽量化の方向に進化している(Zylberberg et al. 2010)。こういった皮骨に見られる軽量化の傾向は浅瀬における運動能力の向上と関連していると考えられている。コズミン鱗はEustenoptelonの後に出現した肉鰭類では消失している。
潮汐域に進出した肉鰭類から四肢類様魚類を経て水棲四肢類まで、Eustenoptelon, Tinirau, Elpistostega, Panderichthys, Tiktaalik, Elginerpeton, Hynerpeton, Ventastega,などが次々に出現した(Ahlberg et al. 2008)(Ahlberg 1999)(Ahlbeg et al. 1994)。20Myという短期間に魚類の形態から四肢類の形態へと進化した(Long & Gordon 2004)。デボン紀末365Myaには両生類とされるIchthyostega, Acanthostega, Tulerpeton が出現している。
潮汐域に進出した肉鰭類の系統では、頭蓋は強度が強化される方向に、頭蓋の扁平化と口吻の伸張化、舌顎骨の縮小化と配置変え、鰓蓋の縮小化から消失の方向に、椎骨と肋骨は部位ごとに異なる形状を取る方向に、鰭は肢の方向に変化している。また、頭骨、椎骨、肩帯、腰帯、四肢、さらには鱗も含めて、皮骨が軟骨に、軟骨が骨に置き換わる方向に進化している。
頭蓋の強化は、構成する各骨がより数を減らし、より大きくなることで達成されている。頭蓋天井の骨のこういった変化は徐々に進んでいるが、脳函はPanderichthysとAcanthostegaの間で急激に変化している。左右の頭頂骨や前頭骨の結合の仕方からAcanthostegaの段階で摂食法が変ったことが示されている(Markey & Marshall 2007)。四肢化と平行して摂食法が変化し、その結果脳函や口周辺といった頭骨の形状に大きな影響を与えたことが分かる。
頭蓋の扁平化は摂食方法と呼吸方法に関係があると思われる。浅瀬や泥濘地を追いかけて、あるいは待伏せて捉える摂食方式になったと考えられ、扁平で開口部の大きな口は機能的であった。また口蓋が広いため空気を飲み込む呼吸法に適していた。
Eustenoptelonの段階で鰭は既に肢に対応した基本的な骨格を持ち、Panderichthysの段階で肩帯と胸鰭は上体を支える程度の機能を獲得していた。一方腰帯と腹鰭の四肢化は遅れており、Tiktaalikの段階になって四肢類の特徴を備えるようになる(Coates et al. 2002)。Acanthostega, Ichthyostega, Tulerpetonといった両生類の出現は、デボン紀後期に15My程度の間をおいて2回起こったとされる大絶滅の最初の一つであるフラスニアン・ファメニアン境界(F/F境界)の大絶滅の後である。鰭の四肢化は陸上歩行のためよりは、植物類が繁茂した浅瀬、あるいは潟における水中生活のためであり、植物類をかき分けたて移動したり、泥濘地を這い回るためとされている。陸棲化が進んだといってもAcanthostegaやIchthyostegaの段階でも水棲が主体の四肢類であった。
初期四肢類は385~365Myaに潮汐域や海岸縁、淡水域、河口といった浅い水域に出現したが、この時期は低酸素の時代であったとされる(Berner 2006)。また、陸上では植物が巨大化して繁栄しており、棲息水圏の富栄養化も加わりしばしば酸素不足状態になったことが予想される(Clack 2006)。こういった環境の変化が陸棲化への選択圧となった可能性がある。鰓はその構造・機能上曝気に対して脆弱であり、水から離れる頻度・時間の増加とともに縮小し、肺機能に頼る必要が増したと考えられる。Tulerpetonの段階でほぼ肺呼吸だけで生活できるようになったとされる。
Long & Gordon(Long & Gordon 2004)は380Mya頃の四肢類様魚類から340Maの陸棲両生類Pederpesまでに、幼体化を含め5段階の主要な変化を経ていると考えた。
380Mya:higher osteichthyan (tetrapodmotph) がデボン紀中期に放散:内鼻孔
372Mya:prototetrapod (Elignerpeton) がフラスニアン世に出現:頭蓋の扁平化
360Mya:水棲四肢類 (Acanthostega) がファメニアン中期の初め頃に出現:四肢形成
358Mya:eutetarpod (Tulerpeton) がデボン紀末に出現:肺呼吸
340Mya:truly terrestrial tetrapod (Pederpes) が石炭紀初期に出現:歩行用四肢
上記年代に若干の違いはあるが、基本的には上記のように5段階を経て陸棲化の方向に向かって体の各所が変化したと考えられる。体の各部が陸棲化に向かって系統的に順序だって進化したのではなく、ある種では腰帯が先行して進化し、他の種では指が先行して進化している。棲息環境や生態系によって進化のパターンが異なっていたのであろう。
初期四肢類に関して淡水棲息説と海水棲息説の議論が為されてきた。従来初期の四肢類が棲息していた場所は、大河が蛇行しており、三日月湖もあり、植物が枝や葉を落とし、動物の死骸が流れ着き、泥で覆われた淡水域であり、こういったところで上陸が始まったとされた。また、ローマーによる「干上がった池から水を求めて移動するために四肢を進化させた」とする仮説は、初期四肢類が淡水域で出現したことを主張している。しかし、最近の調査では、こういった陸棲化への道筋は一本だったわけではなく、生息環境などによって様々の様相があったとされる。完全ではないにせよ上陸を果たした初期両生類は只1種であったわけではない。ほぼ同じ時代365Myaに棲息していたIchthyostegaとAcanthostegaは棲息場所も体型も生活方式も異なっており、既に四肢類が多様化していることをうかがわせる。
陸棲化を可能にした脊椎動物はきわめて短期間に多数の種を形成しており、ユーラメリカ大陸やゴンドワナ大陸の緯度30度以内の地域の淡水域、汽水域、海水域いずれからも化石が出土しており(Blieck et al. 2007)(Goldberg et al. 2005)(Benton 2010)(Benton & Donogue 2007)、さまざまの環境から何種類もが上陸を試みたと考えられる(ジンマー)。
実際この時期に11系統の初期四肢類が存在したとされる(Carroll 2005)。デボン紀に40~50種ほどの四肢類の化石が出ている。いずれも側線と鰓を持ち、水棲中心の生態系に棲息していたことをうかがわせる。指を持つのはAcanthostega, Ichthyostega, Tulerpetonの3種のみであり、指の数も6~8本とまだ定まったものではなかった。
デボン紀末になるとユーラメリカ大陸とゴンドワナ大陸間での四肢類の往来があり、地域特有性が薄れ始めた。この頃に二回目の大絶滅が起こった(Clement et al. 2004)。これら初期四肢類はこの大絶滅事変に遭遇し、多くは生き残れなかった(Carrol 2002)。デボン紀末から石炭紀初期の345Myaまでのおよそ15My間は脊椎動物の化石がほとんど出土しない所謂「ローマーの空隙」といわれる時代である。
現時点では、この空隙を埋める化石としてスコットランドの350Myaの地層から出たPederpes(Clack 2002)とそれより若い346Myaの地層から出たWhatcheeriaの2種が知られているのみである。これらは5指であり、頭骨も扁平でなく幅が狭くなっていおり、石炭紀に繁栄する新たな四肢類の特徴を既に備えている。
初期四肢類とされるAcanthostega, Ichthyostega, Tulerpeton等とは大きく異なった体形を持つ新四肢類は「ローマーの空隙」が終わった直後の石炭紀初期のヴィゼアン期335Myaには急激に放散しており、少なくとも3つのグループ、迷歯亜綱:Labyrinthodontiaの分椎目:Temnospondyliと炭竜目:Anthracosauria、及び空椎亜網:Lepospontyliに分かれるほど多様化している。 新たに出現した四肢類の多くは水棲である。氾濫原や河口域、河川といった淡水環境に棲息する両生類が多かったが、海生の四肢類も存在していた(194 Garcia et al. 2006)。また、陸棲種も増加し始めている(Clack 2001, 2002)。この頃出現した主な四肢類のCasineria、Eucritta、Greererpetonはいずれも小型で5指を持っている。
「ローマーの空隙」は、海生、淡水生の生物のいずれも大絶滅をした直後なので実際に脊椎動物もほとんどいなくなったことを反映していたと考えられた。しかし、第1章の図7で見たように総ての脊椎動物が大絶滅をしたわけでは無いことが明らかになってきた。
また、「ローマーの空隙」終了後すぐに新奇な脊椎動物の急激な出現と放散が見られる。従って、「ローマーの空隙」はほぼ絶滅したデボン紀の初期四肢類のニッチを占めるべく様々の試みが為されていた新四肢類揺籃の時代であったと考えられる(Carrol 2002)。即ち、全体的には脊椎動物の個体数は激減したであろうが、種の数はむしろ増加したと思われる。最近では、「ローマーの空隙」は化石収集が不完全なために見られる現象であり、当時の実際の脊椎動物の生態系を反映していないとする論文が出されている(Smithson et al. 2012)。
図4に肉鰭類・四肢類の分岐図を示す。デボン紀末に急激に四肢類の系統が増加したがごく小数の系統のみが石炭紀まで生き延びている。「ローマーの空隙」の終了直後に冠四肢類が急激に多様化している。
分椎目は迷歯亜網最大の目で、水棲、両性、陸生、海生と多様な環境に適応した。現生両生類とこの頃の両生類とは大きく異なっており、そのため従来から現生両生類は一括して平滑両生亜細網:Lissamphibiaとして別に分類されている。現生両生類が分椎目と空椎亜網のどちらの系統なのか、あるいは両方の系統につながっているかに関する議論は継続中である。最近は分椎類のバラネルペトンが現生両生類の祖先とする見解が出されており(クラック)、有力視されている。バラネルペトンはヴィゼアン期中期333Myaのスコットランドの地層から化石が産出している。側線や鰓弓が無く四肢の骨化も進んでおり、かなりの程度に陸生に適応していたとされる。

From Coates et al. 2008
一方、現生羊膜類は炭竜目:Anthracosauriaのエンボロメリ亜目から生じたとされており、爬虫類に分類され羊膜類の祖先とされるウェストロティアナ:Westlothianaもスコットランドの同じ地域で同じ時代に生きていた。スコットランドから出土した338MyaのCasineriaも羊膜類の特徴を備えており(Paton et al. 1999)、Casineriaやウェストロティアナは羊膜類の幹グループとみなされている。両生類のバラネルペトンと羊膜類のウェストロティアナが同じ時期に同じ地域に生息していたことになる。従って、両生類と爬虫類や哺乳類の祖先種は338Myaよりも早い段階、恐らく「ローマーの空隙」に含まれる360~355Mya頃には分岐したいたとされる(Coates et al. 2008) (Zhang et al. 2005) (Hedges & Kumar 2003)。
多くの初期四肢類がデボン紀末の大絶滅事変で絶滅して以降、恐らく多様な方向で陸棲化が試みられたのが「ローマーの空隙」の時代であったと考えられる。その試みの中で両生類と羊膜類につながる系統が生じたものと思われる。そしてこの新四肢類が現生四肢類へとつながっていくことになる。
第2章-7 ハイギョとシーラカンス
条鰭類と肉鰭類が分岐した直後に肉鰭類の系統からハイギョとシーラカンスが分岐したとされる。我々とより近いのはハイギョ目なのか、総鰭目なのかに関して議論があり(Schultze 1994)(Zardoya et al. 1998) )(Zardoya & Meyer 1997) (Yamanoue et al. 2006)(Zhu & Schultze 1997)(Yokobori et al. 1994)、当初は総鰭目とする見解が有力であった。しかし、分子生物学に基づいた最近の研究ではハイギョ目の方が我々に近いとする説が有力視されているようである(Janvier & Clement 2010) (Maisey 1986)(Ahlberg 1991)(Cloutier & Ahlberg 1996)(Meyer 1995)(Zhu et al. 2001)(Brinkman et al. 2004)(Venkatesh et al. 2001)。しかし、最近ハイギョとシーラカンスは単系統の姉妹群であり、この姉妹群が四肢類に最も近縁とする説が出されている(Shan & Gras 2011)。ハイギョ、シーラカンス、四肢類の間の系統的な関係に関しては依然として議論が続いているようである。
ハイギョの化石は1827年に報告されていたが、生きたハイギョは1833年にアマゾン河で、1840年代になるとアフリカや南米で種々のハイギョが捕獲されている。ダーウィンは古生代と現生のハイギョの類似性、及び解剖学的知見を元にハイギョは進化の移行段階にある「生きた化石」と考え、彼の進化論を補強した。なお、現生ハイギョは3種でありいずれも淡水性であるが、初期のハイギョは海生であった。
ハイギョの系統で最も古いのはシルル紀末に棲息していたStylocithysであり、417Mya頃には出現していた(Zhu & Yu 2002)。
初期ハイギョは、頭蓋における骨間の結合は強化されており、また頑丈な顎骨を持っており、強い咀嚼力があったことを予想させる。脳函は一体化された骨化の進んだ骨からなり、咀嚼のための筋肉が付着する場所の骨は厚みが増している。また、後の系統になるほど口中の体積が大きくなっており、より多くの空気を飲み込むのに適した形態になっている。しかし、内鼻孔は無い。鼻から下顎にかけてsensory-line canals があり、混濁した泥の環境に棲息していたと考えられる。歯はエナメルで覆われている。鰓と肺を持ち、また、ハイギョの心臓の心房は不完全ながら二つに分離されている。皮骨と鱗はコズミンであり、pore-canal systemが見られる。二つの背鰭と非対称な尾鰭を、またひも状の、あるいは未発達にみえる鰭を持つ。
現生ハイギョの循環系は両生類のものに似ているが、初期四肢類出現より前に肉鰭類の系統から分岐しており、従って四肢類とは独立に進化したとされる(Zhu & Alberg 2004)。なお、肺は四肢類と同様消化管の前側から分岐し、左右に2個対になってやや背側に位置している。
シーラカンスは白亜紀末に恐竜類と一緒に絶滅したとされていたが、1938年に東アフリカでその生存が確認された。その後1952年に最初の発見地からかなり離れたマダガスカル島近くのコモロ諸島で、さらには1998年に別種のシーラカンスがインドネシアで捕獲されている。互いに離れた3箇所でシーラカンスが棲息していることは、プレートテクトニクスに基づいた大陸移動によって説明されている(Springer 1999)(Uyeno & Yabumoto 2008)。
肉鰭類の系統からハイギョの祖先とされるStyloichthysは417Myaには分岐しているが、シーラカンスは同じ頃、乃至はハイギョより古い時代に分岐したと考えられる。しかし、分岐直後の化石はあまり多くはない。従来は408Mya頃に生息していたEoactinistiaが最も古いとされていたが(Johanson et al. 2006)、最近418Myaの地層からCrosspterygiiが出土している(Brazeau 2009)(Young 2010)。
現生シーラカンスの主要な形態はデボン紀からあまり変化していず、その進化速度が非常に遅いことで知られている。しかし、最近411Mya頃に棲息していたとされるシーラカンス系の化石Euporosteusが既に現生種化していることが明らかになり、この頃までにほぼ進化を終えていたことになる(Zhu et al. 2012)。即ち、シーラカンスは分岐直後に急激な進化をしたが、その後はほとんど変化しなかったと考えられる(Friedman & Coates 2006)。
シーラカンスは特徴的な管状の脊柱を持っており、こういった形状は脊柱の初期形態と考えられる。シーラカンスが胎生であることは、腹中に小さな魚を伴った化石から推測されていたが、現生種からそれが確認された。頭蓋に関節があることも化石肉鰭類に共通した特徴であるが、内鼻孔を持たないとされている。ほとんど同じ構造を持つ2つの背鰭のほかに尻鰭を余分に持っているのは、進化の過程で突然変異により生じた特有の特徴とされる。
現生シーラカンスの心臓は一心房一心室と言われているが、心房は分離が不完全ながら二つに分かれているともされている。鰾は条鰭類と同様消化管の後ろ側に位置しているが、鰾内には気体の代わりに脂肪が充填されている。このような鰾は、深い海における浮力を確保するための適応とされている。対鰭は海底を這うためではなく泳ぐのに使っていることが確認されている。浸透圧調整には軟骨魚類と同様に体液の尿素濃度を高める方式を採用している。

第2章-8 条鰭類
条鰭類は上陸にかかわる主題から外れるので以下に概略だけを述べるにとどめる。420Myaまでには条鰭類は肉鰭類と分かれたが、その放散の開始は肉鰭類よりやや遅れていたようである。
初期の条鰭類としてLigulalepisの鱗と思われる化石が中国のYunnanの420Mya地層から出ているが、確実ではない。カナダの400Myaの地層から出土したDialipinaは初期硬骨魚という説もあるが(Friedman & Brazeau 2010)、条鰭類とする説の方が強いようである(Janvier 1996)(Zhu et al. 2006)(Friedman 2007)(Botella et al. 2007)(Esbaugh & Tufts 2006)。しかし、400Myaあたりでは条鰭類はまだ本格的な放散はしていない。条鰭類の多様化は393Mya頃から始まっているようであり(Zhu et al. 2009) (Clack 2006)、デボン紀中期になると本格的に放散し、Cheirolepis、Mimia、Gogosardinaなどが出現している。これら条鰭類はいずれも似たような体型をしており、体長も1メートルを超えることはなくほとんどが15センチ程度であり、まだ生態系で優位な位置を占めていなかった。しかし、デボン紀末の370Myaあたりまでには生態系における主要な位置を占めている。
従来の化石解析から、条鰭類は、肉鰭類から分岐した後軟質下網:Chondrosteiを経て、全骨下網、真骨下網と進化してきたとされた。デボン紀初期に軟質類の系統から現れたBrachiopterugiiは現生のポリプテルスの祖先と考えられ、さらに相前後して現生チョウザメにつながる系統が分岐したとされている。軟質類に分類される初期条鰭類は、石炭紀に入ると急激に放散し、ペルム紀には後の全骨魚類:Holosteiへとつながるレドフィエルフィアを生み出した。現生種ガーはペルム紀に軟質類から分岐した末裔とされる。三畳紀初期には棘魚類が絶滅し、三畳紀後期になると軟質類も衰退し、全骨魚類が繁栄する。現生種アミアはこの時期に全骨類から分岐している。三畳紀からジュラ紀にかけて全骨魚類のフォッドフォラスから現生条鰭類の祖先になる真骨魚類:Teleosteiが分岐し、一方この頃に軟質類がほぼ絶滅した。その後全骨魚類は白亜紀までは繁栄したが始新世にはほぼ絶滅し、以降真骨魚類の時代になり、今に到るとされる。
魚類の移動にとって重要な尾鰭の形状は、軟質類では歪尾であり、全骨類も形状は正尾であるが構造は歪尾である。いわゆる正尾は真骨魚類になってから獲得しており、推進機能が格段に向上したと思われる。
最近の遺伝子解析でも細部に違いはあるが、図6に示すように上述の化石の結果とほぼ似たような結果が得られている (Inoue et al. 2005)。なお、最近は全骨亜網としてひとつにまとめる根拠が弱いということで、全骨亜網と真骨亜網を合わせた新鰭亜網いう分類名が用いられているようである。
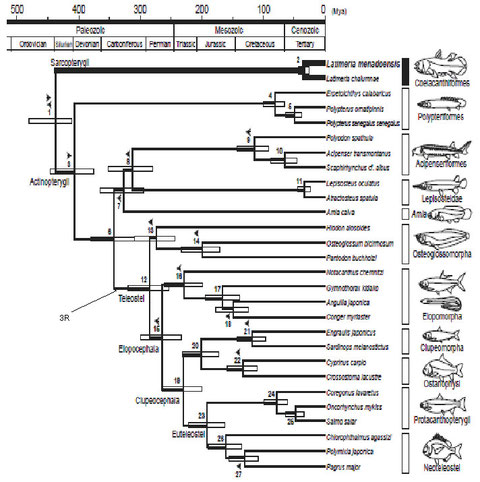
3RはTeleosteiの分枝のところで起こっている From Inoue et al. 2005
ところで肉鰭類との分岐後に条鰭類の系統で、全遺伝子重複が起こったとされる。この全遺伝子重複は、脊椎動物の系統では3回目の全遺伝子重複と考えられfish(or teleost)-specific duplication あるいは簡単に3Rと呼ばれる、
分子生物学的手法により3Rの起こった時期が求められている。しかし、3Rの後に遺伝子の消失や部分的な遺伝子重複などの擾乱があったためその時期の算定は困難である。算定の結果にはかなり幅があり、450Mya~280Myaの間でばらついている (Eichenlaub & Etwiller 2011) (Yan & Cai 2010) (Sato et al 2009)(Kassahn et al. 2009)(Kuraku & Meyer 2009)(Chambers et al. 2009)(Sato & Nishida 2007)(Finn & Kristoffersen 2007)(Semon & Wolfe 2007)(Pavoni et al. 2007)(Venkatesh et al. 2007)(Crow et al. 2006)(Meyer & Van de Peer 2005)(Vandepoele et al. 2004)(Chiu et al. 2004)(Van de Peer 2004)(Hoeg et al. 2004)。
3Rの起こった時期はまだ明確ではないが、こういった3Rに関する一連の研究の結果、現生のポリプテルスやチョウザメはもちろん、全骨下網とされるガー目やアミア目も3Rを経ていないことが明らかにされた。即ち、真骨魚類の系統でのみ3Rが起こっていることになる。
この条鰭類の放散は、条鰭類の系統で起こったとされる全遺伝子重複(3R WGD)と関係がある可能性があるとされる(Kasahara et al. 2007)(Kassahn et al. 2009)。しかし、条鰭類の中でも3RWGDを経ているのは真骨類の系統だけであり、この系統の放散は石炭紀以降に起こったことが化石記録から予想される。
また、WGDが必ずしも多様化を引き起こすわけではないの説もある(Sato & Nishida 2009)。実際に遺伝子解析で得られた図6の魚類の分岐図においても、3RWGD後に急激な多様化は見られず、時代とともに段階的に多様化している。従って現時点では、条鰭類の系統で起こった3RWGDが条鰭類の急激な放散を引き起こし、それが肉鰭類の底生化を促した可能性は低いと考えられる。