 魚が陸上を歩くまで
魚が陸上を歩くまで
第1章 地誌と環境
この章では、主にカンブリア紀後期から石炭紀前期の地球環境と生態系を概観し、その後酸素分圧が進化に及ぼした影響に関する最近の報告をまとめる。
第1章-1 地球環境と生態系
太陽系の、従って地球の誕生はおよそ45億年前(4500Mya)とされる。そして驚いたことに38.5億年前(3850Mya)頃には既に地球上に単細胞生物が出現していた可能性が高いと言われている。その後地球の冷却とともにマントルの粘性が下がり、約27億年前(2700Mya)に二層のマントル対流から一層のマントル対流へと変わったとされる。これにより地球磁場が強くなり、高エネルギー粒子が地表に届かなくなった。その結果DNA損傷の恐れが減り、生物は浅海に進出可能になった。そのため太陽光の利用が可能になり、酸素発生型の光合成をするシアノバクテリアが繁茂し、大気中酸素量が増加し始めた。単細胞同士の共生と相互作用を経て20億年前(2000Mya)頃には真核細胞が出現した。
マントル温度は低下し続けたが、7億5千万年前(750Mya)頃になるとプレートテクトニクスにより潜り込む含水鉱物が深さ30キロの地殻層とマントル層の境界まで含水状態で達することが出来るようになり、それ以降海水がマントル層へ注入され始めた。その結果海水の減少が始まり、大規模な海退現象が起こった。これによりそれまで地球表面の5%程度であった陸地面積が30%あたりまで増加した。また、マントル層への海水の流入によりマントルの粘性が減少し、プレート移動が促進され、550Myaにはゴンドワナ超大陸が形成された(熊沢他2002)(丸山&磯崎1998)。こいうった地表の変化に伴い巨大河川が形成され、陸地からのナトリウム等の流入量が増え海水塩分濃度の急激な増加が始まった。また、河川の発達により堆積岩の形成が促進され、有機物含有堆積物の増加による大気中酸素分圧の急な増加が起こった。
酸素分圧の上昇はオゾン層の形成を促した。さらに酸素分圧の増加により生物は多細胞化と大型化が可能になった。先カンブリア紀の550Mya頃には、多種多様な軟体性の多細胞のエディアカラ生物群が出現した。これらは一般にかなり大型であるが、非常に薄い体をしていた。こういった形状は体を構成する各細胞への酸素を拡散によって供給していたためと考えられる。現生動物の体は一部の種類が放射対称性を持つが、ほとんどは鏡映対称性を持っている。一方、先カンブリア紀に出現した多細胞生物は現生動物に見られない回転対称性や螺旋対称性等特有の対称性を持つ動物もいた。こういった初期の多細胞動物の多くは相互の捕食関係がまだほとんど無く、そのため比較的単純な生態系であったと思われる。
さらに古生代最初期のカンブリア紀になると、530Mya頃に澄江(チンジャン)動物群や505Mya頃にパージェス動物群が出現、急激に多様化した。これらは現生動物と同様鏡映対称性を持ち、多くは数センチメートルと小型である。複雑な器官を発達させた種々の無脊椎動物群の他に脊索動物群も含んでおり、現生の動物群、またはその傍系に関連するものと考えられている(Shu et al. 2003)(Degan 2003)。またその棲息深度は、澄江動物群が水深100~150メートルの大陸棚、パージェス動物群が水深50メートル程度とされ、未熟な移動機能しか持たず、体構造も軟弱であったために暴風時波浪限界ないしはそれより深い水深に棲息していたようである。これらはエディアカラ生物群とは関連性が無いと言われた時期もあったが、最近ではチンジャン動物群、パージェス動物群共にエディアカラ生物群を祖先にしていると考えられている。即ち、先カンブリア紀の動物のうち、鏡映対称性を持つ一部のものと、放射対称性を持つごくわずかのものだけが生き延びたことになる。遺伝子解析によると、脊索動物群の共通祖先は550Mya~564Myaに棲息していたとされる(Putnem et al. 2008)(Hedges & Kumar 2003)。
この後に引き続く脊椎動物の上陸の過程を概観するために、カンブリア紀以降の脊椎動物の歴史を図1に示す。また、古生代が舞台なので古生代の地質年代表を示す(きまぐれ生物学 仲田崇志)。
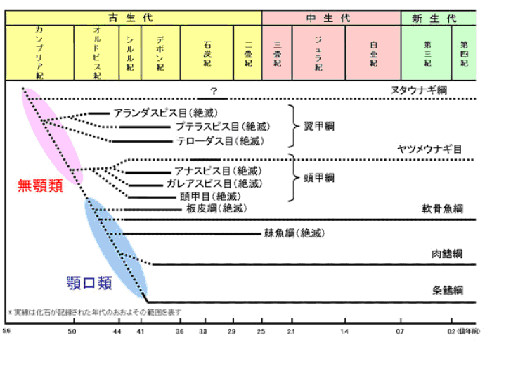
北海道大学COEホームページより
無顎類はカンブリア紀に出現していたが、顎口類はオルドビス紀中期(470Mya)頃に出現したとされる。但し、顎口類の化石記録は鱗を除くと430Mya頃になる(Qu et al. 2010)。顎口類の板皮類、軟骨魚類、棘魚類の系統上の位置関係や出現時期に関しては議論が続いており、確定されていない(Ahlberg 2009)(Janvier 1996).)(Brazeau 2009). (Hallstrom & Janke 2009) (Dosay-Akbulut 2006)。しかし、最初に板皮類が現れ、軟骨魚類、棘魚類が相次いで出現し、やや遅れて初期硬骨魚類が現れ、まもなく初期硬骨魚類から肉鰭類と条鰭類に分岐したとする説が、現在もっとも普通に受け入れられているようである。

オルドビス紀の生態系
カンブリア紀末に大絶滅があったが、オルドビス紀に入ると生態系は急激に回復し、多様化した。従来オルドビス紀は温暖な気候が続いたとされたが、最近ではむしろ海水温度の急激な変化があったと考えられている(Trotter et al. 2008)。即ち、オルドビス紀初期には海水面の温度は40℃を越す高温であったが、オルドビス紀中期までには30℃まで低下しており、この温度低下が急激な生態系の回復を引き起こしたとされる。
また、ゴンドワナ大陸分裂開始による生態系のnichesの増加があった。さらに酸素量の増加によりオゾン層が強化されたため紫外線の影響が減り、浅い汽水域や河口での生存が可能になった。こういったことにより棲息域が拡大されたと考えられる。

From http://www.geocities.co.jp
オルドビス紀の生態系は浅海が主であり、浅海性生物(腕足類、貝、棘皮類、コケムシ類、珊瑚虫類、三葉虫類、筆石類、コノドント類等)が出現・繁栄した。海岸部では470Myaには地衣類(green algae)が上陸、やがて淡水域に進出した。地衣類の淡水域への進出に伴い、河川下流域や汽水域でも生態系が豊かになり、河川、河口、汽水域、浅海域の富栄養化が進んだ。その結果プランクトン量の増加とfilter feeding の動物類が急増した。カンブリア紀には肉食動物として泥中を這う腐肉食動物とごく少数の捕食者がいただけであったが、オルドビス紀になる生態系の拡大とともに肉食性の種類が増えた。こういった環境の下で無顎類が繁栄し、オルドビス紀にはカンブリア紀大爆発にも勝るとされる生物の大放散が起こった。
オルドビス紀の初期480Mya頃には魚類の祖先と考えられる無顎類のアランダスピスが出現していた。しかし、脊椎動物類でもまだ移動性能が低かったため海が障壁となり、オルドビス紀中期までは陸塊毎に異なる生態系を形成していた。
軟骨魚類は、その骨の性質上化石として残りにくいが、軟骨魚類のものと思われる鱗の化石が455Myaの地層から出ており(Sanson, et al. 1996)、オルドビス紀後期には移動性に優れた軟骨魚類が出現し、外洋:open oceansに進出して広範に棲息域を拡大し始めた。また、オルドビス紀後期になると棘魚類や板皮類の化石報告もあり(Long 2011)、遅くとも430Myaまでには板皮類、棘魚類といった顎口類が相次いで出現し、さらに初期硬骨魚類がこれらよりやや遅れて出現したと考えられる。
分子時計に基づいた計算ではカンブリア紀の564Mya(Hedges & Kumar 2003)(Kumer & Hedges 1998)、あるいはオルドビス紀初期の485Mya(Yamanoue et al. 2006)に軟骨魚類が分岐したとされており、また浸透圧調整ホルモンであるナトリウム排出たんぱく質(natriuretic peptides)の比較では528Myaに分岐したことを示している(Inoue et al. 2003)(Blair & Hedges 2005)。こういった計算では概して化石記録よりも古い分岐年代が算定される傾向があるにせよ(Donoghue & Benton 2007)(Hedges & Kumar 2003)、455Myaに既に軟骨魚類が棲息していたとすることは妥当性があると思われる。また、肉鰭類と条鰭類の分岐年代を分子時計に基づいた計算では450Myaに見積もっており、化石記録では450~420Myaとなっている(Benton & Donoghue 2007)(Benton 2005)。
オルドビス紀―シルル紀境界大絶滅
オルドビス紀末の450~440Myaに五大絶滅の一つと云われる規模の大きな絶滅があった。この時期に、ゴンドワナ大陸の南極への移動によりかなり急激に寒冷化・氷河期となり、海退が起こり、海生生物の棲息域である大陸棚が減少した。このため中緯度の浅海域を棲息地とする地域特異性を有していた動物類は、一部はopen oceansに逃れたが、避難地が無い場合には絶滅へと追いやられた。さらにせいぜい50万年ほどの氷河期のあと急激に温暖化が始まったため、open oceans に逃れた種類も多くは絶滅へと追いやられた。100万年を置かず2回起こった急激な環境変化のために、特に中緯度の生態系が大きな影響を被り、浅海性の無顎類やサンゴ、ウミユリなどのオルドビス紀動物群が大きな影響を受け、海生生物の60%以上が絶滅したとされる(Sheehan 2001)(Vandenbroucke et al. 2010)(Trotter et al. 2008)。また、遊泳性があまりなく、地域ごとに適応していた浅海性の板皮類も大絶滅により大きな影響を被ったとされる。
シルル紀の生態系
シルル紀初期には顎口類にとってシルル紀ギャップともいうべき化石の少ない期間がある。シルル紀中期430Myaに棘魚類の(Qu et al. 2010)、デボン紀初期の415Myaに軟骨魚類のPtomacanthusの化石報告(Brazeau 2009)があるが、顎口類のこの頃の化石報告はそれほど多くは無く、個体数の大幅な減少を反映していると思われる。顎口類は、オルドビス生物群とは異なり広範な地域に適応した表現型を獲得するために、その回復・放散は時間を要したと思われる。このため硬骨魚類を含めた顎口類の回復・放散は、シルル紀後期からデボン紀初期になった(Zhao & Zhu 2010)。従ってオルドビス紀末の大絶滅後地球全体の生態系が回復するには、1500万年ほどを要したことになる(Zhao & Zhu 2010)(Sheehan 2001)。
無顎類などのオルドビス紀動物群は移動性があまり無かったが、オルドビス紀末までに既に充分に放散していたと考えられ、特に赤道地帯にあったLaurentia大陸では絶滅を生き延びた種も多かったと考えられる。生き残ったオルドビス生物群は、表現型の大きな変化を必要としなかったため大絶滅事変終焉後比較的単純な生態系から出発し、immigurationが関与して広大なnichesに放散し、500万年程度で急速にほぼオルドビス紀に近い生態系を回復した(Krug & Patzkowsky 2004) (Krichner & Weil 2000)。また、オルドビス紀から引き続いて植物類の陸上への進出があり、シルル紀後期には捕食性の蜘蛛類、多足類も陸に出現した。このため河川、河口、汽水域、浅海域の富栄養化が進み、オルドビス生物群が引き続いて繁栄した。

From http://www.geocities.co.jp
一方軟骨魚類、棘魚類、板皮類といった顎口動物群は出現後まもなくあまり放散しないうちにオルドビス紀末の大絶滅に遭遇したことになる。顎口類は充分な多様化をする前に大絶滅に遭遇したため、大絶滅終焉後はごく少数の種と個体数しか生き残らなかったと考えられる。しかし、遊泳性が劣るため大絶滅の影響を受けたと予想される板皮類でも、大絶滅事変が終焉した500万年後にはAntiarchiの化石報告があり(Qu, et al. 2010)、低緯度地方に生息していたごく一部が大絶滅から逃れることが出来たようである。一方、遊泳性の軟骨魚類や棘魚類は底生性の板皮類よりは被害はひどくなかったと予想されるが、大絶滅後の出土化石は少ない。
シルル紀末期にはLaurentia大陸とBaltica陸塊が衝突しEuramerica大陸が形成され、カレドニア造山活動が盛んになった。Laurentia大陸とBaltica陸塊の衝突によりイアペトウス海が消滅したため、海生生物の棲息域が減少し小規模な絶滅が起こったとされる。小規模とはいえこの絶滅では海洋の高次消費者から1次消費者まで影響が及んでおり、放散虫、筆石、コンドント、アンモナイト、魚類の系統が大幅に入れ替わっている(熊沢他)(丸山&)。
この時期の魚種は必ずしもどの系統に属するのか明確でないものも多く、例えば419Mya頃棲息していたGuiyu (Qiao, & Zhu 2010)は肉鰭類に分類されているが、背鰭の数や形状は板皮類に、脳函は肉鰭類に似た特徴を示している。420Mya頃出現したAndreolepisとLophosteusも中間的な特徴を示しているが、現在では硬骨魚類とされている(Botella et al. 2007)。AndreolepisとLophosteusは肉鰭類と条鰭類の分岐直後の時期に棲息していた硬骨魚類であるが、初期硬骨魚類、乃至はその特徴を強く持つ硬骨魚類と考えられる。また、GuiyuやPsarolepisも肉鰭類に分類されているが、初期硬骨魚類に非常に近いとされる(Zhu & Yu 2009) (Zhu et al. 1999)。
初期硬骨魚類も430Mya頃からあまり遅れない時期に出現したとされるが、体形が分かる化石報告はシルル紀のほとんどの期間にわたり無い。棘魚類や軟骨魚類に対してまだ生態系での位置を確立できず、個体数が少なかったと考えられる。また、初期硬骨魚類は遊泳性であり(Smith et al. 2002)、沿岸から比較的離れた海域を主な生息場所にしていたと予想され、このために化石として残りにくかった可能性もある。最も古いと考えられる初期硬骨魚類の化石は422Myaに棲息していたAndreolepisである(Hedges & Kumar 2003)。
シルル紀中期に棲息していた主な脊椎動物類を図4に示す。初期硬骨魚類と考えられるAndeolepis (11番)は、遊泳性の棘魚類(10番)と同様かなり洗練された魚形をしており、遊泳性に優れていたことを窺わせる。
化石記録は不十分であるが、初期硬骨魚類は420Myaまでには肉鰭類と条鰭類に分岐したとされる(Zhu et al. 2009)。シルル紀最後期の410Myaあたりから肉鰭類は急激に多様化している (Botella et al. 2007)(Sonkoly et al. 2007)(Dosay-Akbulut 2006)。最初期のハイギョとされるPorolepis が416Mya(Zhu et al. 2009)、Styloichthysが414Myaに出現しており(Zhu & Yu 2002)、最初期のシーラカンスとされるEoactinistia が408Myaの化石(Johanson et al 2006)として報告されている。この頃の肉鰭類の多くは軟骨魚類や板皮類、棘魚類の特徴も兼ね備えているが(Zhu et al. 1999)、これら肉鰭類の多くは、その体形から初期硬骨魚類の遊泳性を引き継いでいることを示している。また、これら肉鰭類は非常に洗練された魚形を示しており、410Myaよりかなり以前に既に出現し、適応・放散していたことを予想させる。条鰭類は肉鰭類よりもやや遅れて390Mya頃に放散している(Zhu et al. 2009)。

図中の魚類は総て海生である。無顎類; thelodonts (1:Phlebolesis), heterostracans (2:Toypelepis), anaspids (3:Rhyncholepis), osteostracans (4:procephalaspis, 5:witaaspos, 6:thyestes,7: dartmuthia, 8:trematapis, 9:oeselaspis)と有顎類; acanthodians (10:Nostolepis), osteichthyans (11:Andeolepis)。osteostracans類はラグーンや浅海、thelodontsとacanthodiansは表海水層(epipelagic:水深100m以内の太陽光の届く海表面域)に棲息していた。無顎類のthelodontsは、初期の形は平たく海底生だったが、シルル紀後期には硬骨魚に似た形になった。 From
デボン紀の生態系
デボン紀前期には赤道をまたいでユーラメリカ(Euramerica)大陸があり、その東側から南にかけて間に比較的狭い海をはさんでより大きなゴンドワナ大陸があった。ゴンドワナ大陸の北岸沿いで、またユーラメリカ大陸とゴンドワナ大陸の間でも脊椎動物の移動・交雑があったことが、化石記録から結論される(Zhao & Zhu 2010)。
デボン紀前期から中期にかけて陸上の植物は枝分かれしたより複雑な形態を持つようになり、高さも2メートルに達するものも出てきた。維管束植物の繁栄によりデボン紀中期・後期にかけてさらに河口域や淡水域に豊かな生態系が形成された。デボン紀後期には樹高20メートルにも達するトクサ類、シダ類からなる大森林が形成され、さらには種子を持つ植物が出現し乾燥地にも進出した。また、森林にはムカデ類、サソリ類、ダニ類、クモ類、さらにはある種の昆虫類といった節足動物が生息していた。しかし、植物食の節足類はまだほとんど出現していず、肉食であり互いに捕食しあっていた。
デボン紀は前期から中期にかけて温暖であり、陸生植物の繁栄によりユーラメリカ大陸の広大な浅い湾、三角州、入り江に広範な泥の生態系が形成され、水底の無脊椎動物類を捕食するbottom feeding predators の板皮類が繁栄した。また、浅海域からopen oceansにかけて、プランクトン類を捕食する遊泳型の棘魚類が、またそれらを捕食する軟骨魚類や硬骨魚類が繁栄した。シルル紀の無顎甲皮類や腹足類、二枚貝類、オーム貝類は、デボン紀においても大きな変化も無く繁栄したが、アンモナイトには形態上の変化が見られ、三葉虫の多くは減少した。三葉虫の減少は顎を持った魚類や頭足類の出現と関係があるとされる。

From http://www.geocities.co.jp
シルル紀からデボン紀にかけての初期硬骨魚類、肉鰭類、条鰭類の分岐時期、放散時期を図6に示す。肉鰭類は420~400Myaにかけて放散し、条鰭類は遅れて390Mya頃から放散を開始している。

顎口類23系統について153個の形態上の特徴を基にthe most parsimonious tree を作成
地質年代は Gradstein, F. M. et al. A Geologic Time Scale 2004 (Cambridge Uni. Press, 2004) による。 From Zhu et al. 2009
デボン紀中期から後期にかけてユーラメリカ大陸とゴンドワナ大陸が合体し始め、パンゲア超大陸の形成が始まり、カレドニア造山運動が活発化した。フラスニアン期末には年輪を示す樹木が見られ、また落葉樹も出現し、陸上では気候の季節変動が大きくなったと考えられる。この時期は酸素分圧の急激な低下が起こったとされ(Berner 2006)、 山火事は起こりにくかったと思われるが、落葉樹の出現に伴い有機堆積物が増え乾燥地では火事が頻繁に起こり、生態系に影響を与えたとされる(クラック)。
390Mya頃になると遊泳力の優れた条鰭類が放散を始めた。このため遊泳性の棘魚類や肉鰭類は、前者は河川など淡水域に進出し、後者の多くは底生となり、各々条鰭類と棲み分けたと考えられる。
底生になった肉鰭類の中からさらに三角州や潟などの潮汐域へと進出し、四肢類様魚類を経て四肢類へと進化する系統が生まれることになる。四肢類につながるもっとも初期の四肢類様魚類とされているのはデボン紀初期の395Mya頃に棲息していたケニクティス:Kenichhysであり、初期段階の内鼻孔を備えている(Zhu & Ahlberg 2004)。四肢類様魚類はオステオレピス:Osteolepis、ユーステノプテロン:Eusthenopteron等を経て、フラスニアン期前期380Mya頃にパンデリクティス:Panderichtysやエルピストステガ:Elpostostega、ティクターリク:Tiktaalikなどが各所に相次いで出現した。
そのわずか5M~10My後のフラスニアン期後半からファメニアン期には両生類に分類される水棲四肢類のエルギネルペトン:Elignerpetonが372Myaに、さらにヴェンタステガ:Ventastegaが(Zhu et al. 2009)、そして365Mya頃にはアカントステガ:Acanthostega、イクチオステガ:Ichthyostega、そしてチュレルペトン:Tulerpetonが相次いで潟湖、汽水域、淡水域と様々の地域に出現する(Metscher et al. 2005)(Clement et al. 2004)。フラスニアン期からファメニアン期に起こった魚類様四肢類から両生類への変遷は非常に急激であった(Ruta et al. 2006)。生態系が豊富で多様性があり、干満の影響、曝気といった環境ストレスによる淘汰圧が強かったためと考えられる。また、フラスニアン期からファメニアン期にかけて酸素分圧が急激に低下したとされており(Berner 2006)、これも陸棲化を促した要因となったであろう。
カナダのエスクナミック湾化石群は、デボン紀後期のフラスニアン期における海生生態系の多様な様相を示している。板皮類、棘魚類、原始的な条鰭類ケイロレピス、肉鰭類のハイギョ、シーラカンス、ポロレピス、オステオレピス類(特にユーステノプテロン)など様々な化石が見られる。また、四肢類に近いと思われるエルピストステゲや、四肢類と思われる化石としてメタクシグナトウス(フラスニアン・ファメニアン境界)、オブルチェヴィクティス(フラスニアン期)、エルギネルペトン(フラスニアン後期)が出土している(クラック)。
四肢類はデボン紀後期には広範な地域に生息していたが、地域ごとに特殊化している(Clement et al. 2004)。しかし、この時期に登場した四肢類のほとんどはデボン紀・石炭紀境界における大絶滅事変を生き延びられなかった。
F/F境界(the Frasnian-Famennian boundary)とデボン紀末の大絶滅
およそ375Myaのデボン紀後期に五大絶滅の一つとされる大絶滅が起こった。Gondwana大陸の一部に氷河が形成され、寒冷化と海水準低下があった。Euramerica大陸の南部も寒冷化し、加えて海洋における酸欠現象も見られた。
しかし、大絶滅の期間や回数はまだ確定的でないし、さらには原因も多くの説があり、結論は出ていない。大絶滅には2つのピークがあったとする説、300万年くらいの周期で何回か繰り返されたとする説、20~25Myの間に8~10回程度の絶滅があったとする説などが出されている。また、原因に関しても大陸合体の進行により低緯度の温暖な海流が止められたために大陸西側が寒冷化し氷河が形成されたとする説、維管植物の繁栄により大気中炭酸ガスが減って寒冷化したとする説、また水圏の富栄養化により無酸素状態が生じたとする説などが提案されている。
この大絶滅が2つからなるとするのが最も一般的に受け入れられている。最初の絶滅は374MyaのF/F境界(Frasnian-Famennian boundary)で起こったもので、Kellwasser event と呼ばれる。二番目の絶滅は359Myaのデボン紀と石炭紀の境界(Devonian-Carboniferous boundary)で起こったもので、Hangenberg eventと呼ばれる 。かってはKellwasser eventが主であり、Hangenberg eventはKellwasser eventの70%程度の規模だったとされた。
その後の研究ではKellwasser eventでは温暖な浅海性の珊瑚、三葉虫、腕足類、アンモナイトといった無脊椎動物が大きな影響を受けた。一方、Hangenberg eventも低緯度地域で影響が大きかったが、特に脊椎動物への影響が大きく、肉鰭類や四肢類の多くは海生、淡水生の区別無く絶滅している(Sallan & Coates 2010)。
絶滅前に繁栄していた板皮類、棘魚類、肉鰭類の絶滅後の回復はほとんど無く、無顎類や三葉虫類もほとんどが絶滅した。大絶滅後の回復は次の石炭紀になるが、それまでの360~345Myaの1500万年間は脊椎動物の化石がほとんど出ない、いわゆる「ローマーの空隙」と呼ばれる時代になる。
大きな環境変化があった後なので実際に脊椎動物の個体数は大幅に減少したと考えられるが、後述するように360~345Myaの期間に四肢類の多様化が進行していたと考えられ、脊椎動物の種類が少なかった訳ではないと思われる。むしろこの環境変化が脊椎動物類の多様化と本格的上陸を促したと思われる。
さらにはデボン紀末の2つの大絶滅で顎口類の総てが絶滅したり、衰退したわけではないことが明らかにされた(Sallan & Coates 2010)。図7に2大絶滅前後における種数変化を示す。板皮類はKellwasser event 以降急激に衰退し、デボン紀末にはほぼ絶滅した。肉鰭類と四肢類はHangenberg event以降減少している。棘魚類はKellwasser eventの以前から既に減少傾向を示している。他方、軟骨魚類と条鰭類はKellwasser event以降むしろ種類が増加している。

Tetrapoda:Rhizodontidaを除いた冠Tetrapoda(Coates et al. ‘08の分岐に基づく)であり、elpistostegaliansを含む From Sallan & Coates 2010
「ローマーの空隙」における脊椎動物の化石は2箇所からしか得られていず、かなり限定されている。そのため最近Clackのグループは、「ローマーの空隙」は化石収集が不完全なことによるためであり、当時の脊椎動物相の実体を反映していないために生じたものであると結論している(Smithson et al. 2012)。
石炭紀前期の生態系
石炭紀前期はミシシッピ紀ともいわれ、359Myaに始まり345Myaで終わるトウルネージアン世と345Myaに始まり328Myaで終わるヴィゼアン世、328Myaに始まり318Myaで終わるサープコビアン世に分けられている。石炭紀はユーラメリカ大陸とゴンドワナ大陸の間の海が狭まり、現在のアパラチア山脈、アトラス山脈、アルプス山脈が隆起形成され、全般的に火山活動が活発であった。

From http://www.geocities.co.jp
石炭紀の気候は徐々に温暖で湿潤になり、世界的に熱帯性の高温な気候となり,季節性が乏しく、大陸内部に浸入した浅いテチス海が広がっていた。ゴンドワナ大陸の南部は依然として氷河に覆われていたので中緯度から高緯度にかけての生態系は貧弱であった。赤道地域全体に広範な森林が形成されていたが、デボン紀の地表を覆っていた樹木はトウルネージアン世の間に絶滅し、種類の異なる木々と交代した。石炭紀の多くの植物は現在の熱帯性の植物とよく似ていて,年輪形成が見られないものが多い。最初は樹高が2メートルに満たないソテツ類やシダ類の低潅木が多く、現在の針葉樹と類縁の裸子植物や種子植物も含まれていたが、ヴィゼアン世になると次第に湿地帯、乾燥地、高地といった環境ごとに棲み分けがはっきりした植物相になった。また、石炭紀は全般的には温暖であったが、何回か寒冷な時期があり海退と海進が繰り返された。
最初の両生類が現れたのが、デボン紀末のフラスニアン期後期からファメン期前期(380~370Mya)とされるが、F/F境界、デボン紀/石炭紀境界の大絶滅により上陸を開始した初期の両生類のほとんどは絶滅した。いわゆる「ローマーの空隙」は360Myaから345Myaの1500万年ほど続いたが、この期間の355Mya頃には現生種につながる新四肢類の両生類と羊膜類が分岐したとされる(Zhang et al. 2005)。両生類は350Myaまでに主要な10系統を輩出していた(ジンマー)。羊膜類の初期系統とされるトウラーペトンは生存期間が短かったが、338Mya頃に羊膜類と考えられるウェストロシアーナが出現した。ヴィゼアン期末の330Mya頃までには現生する羊膜類の主な系統が確立したとされ、30種ほどの新四肢類が知られている。「ローマーの空隙」の期間からミシシッピ紀までの間は、これら新四肢類の系統で急激な進化が起こった時期である(Ruta et al. 2006)。
先述のように「ローマーの空隙」前後で四肢類はまったく異なっている(Clack 2006)。デボン紀の四肢類は大型で頭部が扁平であるが、石炭紀に出現した新四肢類は初期のヴィゼアン期では小型種が多く、アカントステガ(Acanthostega)、イクチオステガ(Ichthyostega)といった祖先との密接なつながりは見られない(Coates et al. 2008)。デボン紀末に生存していた四肢類11系統のうちただ1系統のみが新四肢類へとつながっているとされる(クラック)。これら新四肢類はヴィゼアン期には既にトカゲ様やウナギ様など多様な形態を示しており、水棲種が多い。「ローマーの空隙」が終了した直後で既に新四肢類が多様化を示しており、従って、新四肢類は「ローマーの空隙」と呼ばれる期間に既に出現し、放散を開始していたことを示唆している。
デボン紀と石炭紀の主な肉鰭類、四肢類様、および四肢類の模式図を図8に示す。多くの四肢類が石炭紀に水棲に戻っていることがうかがえる。新四肢類は初期には小型種が多かったが、石炭紀末までには大型種を輩出している。

(a) デボン紀の四肢類:総て幹四肢類である。1. Gooloogongia, rhizodont. 2. Osteolepis, osteolepidid. 3. Kobaralepis, osteolepidid. 4. Canowindra, osteolopidid. 5. Eusthenopteron, tristihopterid. 6. Tristichopterus, tristichopterid. 7. Gyroptychius agassizi, osteolopidid. 8. Gyroptychius dolichotatus, osteolopidid. 9. Cabonicchthys, tristichopterid. 10. Mandageria, tristichoperid. 11. Eusthenodon, tristichopterid. 12. Glyptopomus, osteolopidid. 13. Tiktaalik, elpostostegalid. 14. Panderichthys, elpostostegalid. 15. Ichthyostega, limbed stem tetrapod. 16. Acanthostega, limbed stem tetrapod. (b) 石炭紀の四肢類:幹四肢類 1~5, 9, 11、冠四肢類 6~8, 10, 12~15. 1. Strepsodus, rhizodont. 2. Megalichthys, megalichthyid. 3. Rhizodopis, megalichthyid. 4. Megalocephalus, megalichthyid. 5. Crassigyrinus, stem tetrapod. 6. Palaeomolgopis, adelospondyl (stem amniote or stem tetrapod). 7. Brachydectes, lysorophid (stem amniote). 8. Urocordylus, nectridean (stem amniote). 9. Greererpeton, colosteid (stem tetrapod). 10. Proterogyrinus, embolomere (stem tetrapod). 11. Pderpes, whatcheeriid (stem tetrapod). 12. Westlothiana, stem amniote. 13. Silvanerpeton, embolomete (stem amniote). 14. Dendrerpeton, temospondyl (stem lissamphibian). 15. Gephyrostegus, gephyrostegid (stem amiote). From Coates et al. 2008
そして320Mya頃には冠羊膜類に分類されるいくつかの四肢類が出現、その中から310Myaには最初の哺乳類型爬虫類である盤竜目アーケオシリスが、305Myaには現生爬虫類の祖先とされる双弓類が、303Myaには最初の植物食の陸棲脊椎動物とされる哺乳類型爬虫類のエダフォサウルスが出現している。なお、次の中生代に大繁栄する双弓類は出現後3500万年間ほどはあまり放散せず、放散を開始してまもなくにペルム紀末の大絶滅を迎えたようである。
第1章-2 酸素分圧変化と進化
脊椎動物の一部がなぜ陸棲化したのか、あるいは何ゆえ肺を、または四肢を獲得したのかに関しては多くの仮説が出されてきた。
陸上を目指した原因に関して幾つか提案されているが、捕食・被捕食の関係からの仮説が多いようである。新たな食料源を求めて、あるいは逆に捕食者から逃れるために、といった説が提案されてきた。デボン紀後期の陸上には昆虫やクモ、多足類などの節足類や蠕形(ゼンケイ)動物類が既に繁栄していたが、アカントステガやイクチオステガといった初期段階の四肢機能では捕獲困難であり、小型過ぎて摂食効率も悪かったであろう。また、最初は捕食者から逃れるために潟や三角州といった浅い潮汐域に進出した可能性はあるが、デボン紀後期の潮汐域ではアカントステガやイクチオステガはむしろ上位の捕食者であったと思われ、陸に逃げ込む必要はそれほど大きくなかったと思われる。
肺の獲得に関して最もよく知られており、長いこと支持され、専門誌でもしばしば引用されているのは、棲息していた池や河がしばしば干上がったことが原因であるとする所謂「涸れ池」仮説である。二十世紀初頭の時点ではデボン紀の化石の多くは赤色砂岩層(Old Red Sandstone)から出土していた。赤色砂岩層は泥の中に含まれる鉄分が空気に曝されて酸化したために形成されたものであり、乾燥した環境で形成されると考えられた。しかし、その後赤色砂岩層は必ずしも乾燥気候や淡水域で堆積するとは限らないことが明らかになり、「涸れ池仮説」に疑問が持たれた(クラック)。
また、四肢獲得に関してもいくつかの仮説が提案されているが、長い間信じられてきたのが、「涸れ池」から水を求めて移動するために鰭が四肢化したというローマーによる仮説である。この仮説は「涸れ池」ばかりでなく、「酸素欠乏」や「過剰個体数」などの悪化した棲息環境か逃れるために四肢が形成されたと解釈することも可能である。ローマーの仮説に基づくと、385Mya頃棲息していた四肢様淡水性魚類のユーステノプテロンの対鰭は四肢と似た骨構造を持っており、この鰭は旱魃時には陸上を水を求めて移動するために、そして普段は水底を移動するのに四肢と同じように使われたと考えられた。しかし、現生シーラカンスは鰭を水底移動に使っていないことが明らかにされた。
何ゆえ脊椎動物が陸棲化したのかに関しては、上述以外にも諸説が提案されてきたが、
明確になっていない。最近カンブリア紀以降現在までの大気中酸素分圧の変動に関する詳細な計算に基づく報告が為され(Berner & Canfield 1999)(Berner 2006)(Bergman et al. 2004)、その結果を元に酸素分圧と生物進化を関連付ける論文が増えている(Geroge & Blieck 2011)(Dahl et al. 2010)(Butterfield 2011)(Dahl et al. 2011) (Qu et al. 2010)(Ward et al. 2006)(Clack 2007)(Clement & Long 2010)(Dudley 2000, 1998)(Lenton 2002)(Harrion et al. 2010)(Igamberdiev & Lea 2006)(Benton 2010)。
酸素分圧の変遷
大気の酸素分圧は、1500Myaから700Myaあたりまでは2%程度であるが、700Mya以降急激に増加し始めている(Shikazono 2008)(Payner et al. 2009)(Sessions et al. 2009)(Kirschvink and Kopp 2008)(Catling et al. 2007)(Canfield 2005)(Lenton 2002)。顕生代以降の大気中酸素分圧に関しても幾つかの報告がある (Berner 1999, 2000, 2003, 2006)(Bergman et al. 2004)(Ward et al. 2006)(Igamberdiev & Lea 2006)(Rouxel et al. 2005)。図10にBerner とBergman による3つの計算結果を示す。

酸素分圧の変動に関するいくつかのモデル計算結果:実線(Berner & Canfield 1999), 点線(Berner 2006), 断続線(Bergman et al. 2004) From Harrison et al. 2010
結果にばらつきはあるが、400Mya以降現在に到るまでの範囲では3つとも大体似たような傾向を示している。いずれもデボン紀中期頃は低酸素分圧であり、石炭紀からペルム紀にかけて酸素分圧が高かったことを示している。また400Myaより古い時代では、Bergman の報告のみが低い酸素分圧を示している。計算に際して入力情報に最新の炭素濃度と硫黄濃度の変化に関する地質学的データを使用していることもあり、酸素分圧と進化の関連性に言及している最近の報告の多くはBerner 2006の結果を採用しているようである。
即ち、肉鰭類が出現し、上陸を始めるまでの間における酸素分圧は420Mya頃に25%程度に高くなった後減少し、380Mya頃に12%程度と最低値になり、その後次第に上昇したが、デボン紀末の360Myaでもまだ18%程度であった。現在と同程度の酸素分圧になるのは石炭紀になってからである。
鉄鉱物は酸化的雰囲気で形成されるので、深海堆積物中に含まれる鉄イオンの詳細分析から堆積当時の酸化還元状態を調べることが出来る。その結果ペルム紀末とジュラ紀前半に酸素分圧が低かったこと、デボン紀後半にも酸素分圧が低くなった可能性があることが明らかになった(Sato et al. 2009)。また、シルル紀末の420Myaにもっとも古い山火事の記録があり、酸素分圧が13%以上になると山火事の発生が可能になることからシルル紀末には酸素分圧が相応に高かったことが明らかにされた(Scott & Glasspool 2006)。これらの結果はBerner 2006の酸素分圧の計算結果とかなりよい一致を示している。こういった傍証からもBerner 2006 の計算結果が現時点では最も信頼性があると考えられている。
酸素分圧変遷と進化
酸素分圧と生物進化を関連付ける最近の論文を以下にまとめる。
酸素分圧と進化の関係を考察した論文は各々注目している現象と時代に違いはあるが、酸素分圧が上昇している時期に新たな動物群が出現しているとしている点では一致しているようである(Dahl et al. 2010) (Geroge, & Blieck 2011) (Ward et al. 2006)(Clement & Long 2010)。エディアカラ動物群は560~550Myaに出現したが、これは高酸素時代であったとされる(Dahl et al. 2010)。また、図6に示すように、オルドビス紀中期の472~461Myaにかけて無顎類の放散が見られるが、これも高酸素時代である。450~420Myaにかけて急に酸素分圧が上昇しており、この時期に顎口類が出現し、423Mya頃には肉鰭類と条鰭類が分岐し、416Myaまでには世界各所に顎口類が棲息するようになった(Qu et al. 2010)。酸素分圧は410Mya頃には25%程度まで上昇したが、この頃に多量の酸素を必要とする大型の捕食性魚類が出現している(Geroge, & Blieck 2011) (Dahl et al. 2010)。また、酸素分圧の上昇に伴い種の数も増えるが、体のサイズも大型化するという一般的な傾向がある(Harrison et al. 2010) (Geroge, & Blieck, : 2011)。
1.無顎類、顎口類の放散と酸素分圧
化石記録でみる限りデボン紀における幹四肢類の出現は375Mya~365Myaであり、この時期は急激な酸素分圧の低下後再び上昇し始めた時期である。海岸や河口の泥濘地に進出していた肉鰭類の一部は酸素分圧の低下に伴い既に獲得していた肺の機能を高める方向に、従って陸棲化の方向に適応した可能性が考えられよう。

2つの灰色で示した期間のうち年代の古い方はオルドビス紀における無顎類の放散の時期であり、シルル紀の灰色部は初期顎口類の放散の時期を示す。これら2つの放散には、海水面温度変化との関連性は見られない。いずれの放散も酸素分圧が上昇中に起こっている。
時間軸は左から右 From Qu 2010
2.陸上植物の繁栄と四肢類の出現
これに対して低酸素化と四肢類の出現は同期しているが、大気中の酸素分圧が低くなったことと四肢類の出現とは直接の関係が無いとする説が出されている(Clack 2007)。酸素分圧の低下に伴い陸生植物の幹の太さが増大している。これは低酸素化に伴い植物が繁茂したことを示しており、そのため浅海や淡水といった水圏の富栄養化が進み、酸素欠乏状態になりやすかったと類推される。391MyaあたりからHangenburug eventにかけて黒色頁岩が増えているのはそういった酸素欠乏が各所で起こっていたためである。酸素欠乏状態に置かれた肉鰭類は既に獲得していた肺に口から空気を飲み込んで呼吸を行う方式を洗練させたであろう。従ってこの仮説は陸棲化に対する「涸れ池仮説」の後を引き継いだ「酸素欠乏仮説」の考えを踏襲していると言える。
2つの灰色で示した期間のうち年代の古い方はオルドビス紀における無顎類の放散の時期であり、シルル紀の灰色部は初期顎口類の放散の時期を示す。これら2つの放散には、海水面温度変化との関連性は見られない。いずれの放散も酸素分圧が上昇中に起こっている。
時間軸は左から右 From Qu 2010
2.陸上植物の繁栄と四肢類の出現
これに対して低酸素化と四肢類の出現は同期しているが、大気中の酸素分圧が低くなったことと四肢類の出現とは直接の関係が無いとする説が出されている(Clack 2007)。酸素分圧の低下に伴い陸生植物の幹の太さが増大している。これは低酸素化に伴い植物が繁茂したことを示しており、そのため浅海や淡水といった水圏の富栄養化が進み、酸素欠乏状態になりやすかったと類推される。391MyaあたりからHangenburug eventにかけて黒色頁岩が増えているのはそういった酸素欠乏が各所で起こっていたためである。酸素欠乏状態に置かれた肉鰭類は既に獲得していた肺に口から空気を飲み込んで呼吸を行う方式を洗練させたであろう。従ってこの仮説は陸棲化に対する「涸れ池仮説」の後を引き継いだ「酸素欠乏仮説」の考えを踏襲していると言える。

網がけ部はジベーティアン-フラスニアン期であり、最も酸素濃度が低いときに対応している。植物の幹直径が急激に増大し、幹四肢類の放散が急激に起こっている。二酸化炭素濃度はこれらの変化とは無関係である。時間軸は下から上 From Clack 2007
3.四肢類の放散と酸素分圧
最近397Myaに指を持った四肢類が生存していたことを示す足跡化石(Niedzwiedzki et al. 2010)が発見され、四肢類の出現はかなり早いことが示唆された。これを説明するために四肢類の放散は酸素分圧最大の時期であるとする説が提案された(Geroge & Blieck 2011)。図13にその結果を示す。青い部分が酸素分圧の高い時期であり、急激に四肢類の種類が増加している。397Myaの足跡もこの高酸素分圧の期間に含まれている。さらには1986年に報告された397Nyaよりも古いデボン紀初期の足跡(Glensla trackway)とされる化石も初期四肢類が酸素分圧上昇の期間に出現したことを支持するとしている。

初期四肢類の系統図:V-Valentia Islandにおける四肢類の痕跡化石 Z―Zachelimieにおける四肢類の足跡化石 G―Glenslaにおける四肢類の痕跡化石
時間軸は右から左 from Geroge & Blieck 2011
4.節足動物、脊椎動物の陸棲化と酸素分圧
また、酸素分圧の低下と「ローマーの空隙」を関係付けた考察も為されている(Ward et al. 2006)。水棲動物の陸棲化には2段階があり、最初の425~385Myaには多足類、クモ類、昆虫類といった節足動物の陸棲化が、385~360Myaには脊椎動物の陸棲化が起こった。陸棲化は、小型動物や脊椎動物のいずれにおいても酸素分圧が低下した時期に中断され、これが「ローマーの空隙」に対応するとしている。酸素分圧の回復と共に再び陸棲化が進行した。図14に四肢類と多足類の例を示すが、「ローマーの空隙」は酸素分圧が最小になった時期よりも3000万年ほど遅れて始まっており、当該主張と合わないようである。

from Ward et al. 2006
5.化石記録に見る陸棲化と酸素分圧の関係
図10のBerner 2006の結果を見ると酸素分圧は408Mya頃に25%になった後急激に減少し、380Mya頃には14%以下まで低下している。3000万年程度で酸素分圧が10%ほども低下したことになり、これが実際に起こったとすれば生物への影響は大きかったと予想される。
酸素分圧が急激に低下したとされる410~380Myaの時期に肉鰭類の系統において多様化に引き続いてコズミン鱗の消失、底生化と内鼻孔の形成、浅瀬への進出と鰭から肢への移行、といった後の陸棲化へとつながる一連の変化が始まっていることが化石から確認されている。そして実際に曲がりなりにも陸に上がれるようになったのは、酸素分圧が上昇し始めた370~365Mya頃である。石炭紀における脊椎動物の完全な陸棲化と放散も酸素分圧が上昇中に起こっている。
計算で得られた酸素分圧に関する留意
以上に見てきたようにBerner2006の酸素分圧に関する計算結果を基に適応・放散や脊椎動物の陸棲化といった進化上の現象の説明が為されている。しかし、Berner2006の計算結果、あるいは酸素分圧の変遷に関わる同様の計算結果を進化の現象と結び付けるには、注意が必要である。
Bergmanの計算結果ではオルドビス紀からデボン紀にかけては低酸素時代であった。一方Bermerは、シルル紀に山火事があったことから、シルル紀が低酸素時代ではなかったとし、Bergmanの計算結果を否定的にみている。しかし、山火事に関するより詳細な調査結果では、350~340Myaの山火事はBernerの結果と一致するが、235~65Myaの山火事はBergmanの結果の方が説明出来るようである(Belcher et al. 2010)。
モデルを基に酸素分圧の変遷を計算したいくつかの報告があるが、いずれも計算に際してC, S, Mo, Fe等の同位体に関する地質学的データを入力している。しかし、これら同位体のデータは、時代が同じであっても分析対象の岩石が堆積した環境や堆積後の来歴によって異なることもあり、非常にばらつきが大きい(Dahl et al. 2010)(Parnell et al. 2010)。さらにBerner2006は、C同位体の結果が直接酸素分圧の計算結果に反映するとしているが、670Mya~現在までをカバーするために計算に用いたC同位体のデータとして20件ほどの論文から引用している(Kirschvink & Raub 2003)。ばらつきの大きなデータをこの期間に渡って繋ぎ合わせるには人為的な操作を入れざるを得ず、恣意性が入り込む可能性がある。
Berner2006の計算結果を含めこれら計算結果は一般に大きな誤差範囲を見込んでいる(Lenton 2002)。また、古生代が始まる直前に2乃至3回の全球凍結があり、さらに海における酸欠状態があったため、この頃の酸素分圧を算定することは困難とされる(Canfield 2005)。古生代が始まる直前の酸素分圧の算定結果が不確実であれば、この算定結果を酸素分圧の初期値とする古生代以降の計算に不確実さが入り込むことになる。一般に初期値は計算結果全体に大きな影響を及ぼすと考えられる重要なパラメーターである。
さらに使うモデルによって、あるいは考慮する酸素増減に関与する地球物理学的現象の解釈の違いによって計算結果が異なってくる(Holland 2006)(Catling et al. 2007)。また、Berner2006の場合入力する初期値パラメーターが19個もある。Bernerはこれらパラメーターの計算結果への影響度を検討しており、問題は無いとしている。他のモデルでも同程度の数の初期値を使っている。しかし、理論計算においては一般的には入力パラメーターの数が3~4個もあれば、計算結果をかなり変えることが可能である。
現時点では、使用している地質学的データやパラメーターの検討、傍証などから、少なくとも古生代の酸素分圧に関してはBerner2006の計算結果が最も妥当性があるとされているが、こういったことから、酸素分圧の計算結果を進化の現象と結び付けることは注意を要すると思われる。
オゾン層の形成
生物の陸棲化にあたり紫外線が相応に弱くなっている、即ち、オゾン層の形成が必要である。しかし、オゾン層形成時期に関しては光合成細胞が出現した2000Myaころとする説から浅海に無脊椎動物類が進出した450Myaころとする説など諸説ある(Cockell & Raven 2007)。
紫外線からDNAを保護するオゾン層を形成するためには酸素分圧が現在値の5×10-3程度あればよいとされる(Cockell & Horneck 2001)。この説に基づくと酸素分圧に関する予測値から(Lenton 2002)(Holland 2006)、2000Mya頃には相応のオゾン層が形成されていたことになる。
一方、オゾン層が無くても水深30メートルより深ければDNAは安全に保護される(Cockell 2000)。515~505Mya頃棲息していたパージェス動物群は水深50メートル程度の暴風時波浪限界ないしはそれより深い水深に棲息していたが、この深度は紫外線からDNAを保護することにも役立っていたであろう。従って、地表近傍の紫外線強度が生物の棲息にとって安全になったのはこれよりも後の可能性も考えられる。
酸素分圧は石炭紀にはかなり高い30%台に、一方ペルム紀からジュラ紀にかけて15%程度に低くなったとされる。大気中酸素分圧とオゾン層形成の関係が不明であるが、ペルム紀やジュラ紀における陸上生物の繁栄状態から、酸素分圧が15%程度でも生態系は紫外線に対して充分保全されていたと考えられる。酸素分圧は2000Mya以降古生代に到るまで基本的には増加しているが、恐らく酸素分圧が急激に上昇し始める632Myaからカンブリア紀直前の551Myaまでの間に(Sahoo et al. 2012)、紫外線に対して機能的に現在とそれほど違わないオゾン層が形成されたと思われる。